Cell Differentiation and Variation
Stem Cells and Potency
Stem cells are unspecialised cells which have the ability to become specialised cells, such as heart cells or neurons. The process by which a stem cell is converted from an unspecialised cell to a specialised cell is called cell differentiation. Stem cells have an unlimited capacity to divide and can produce lots more stem cells by mitosis.
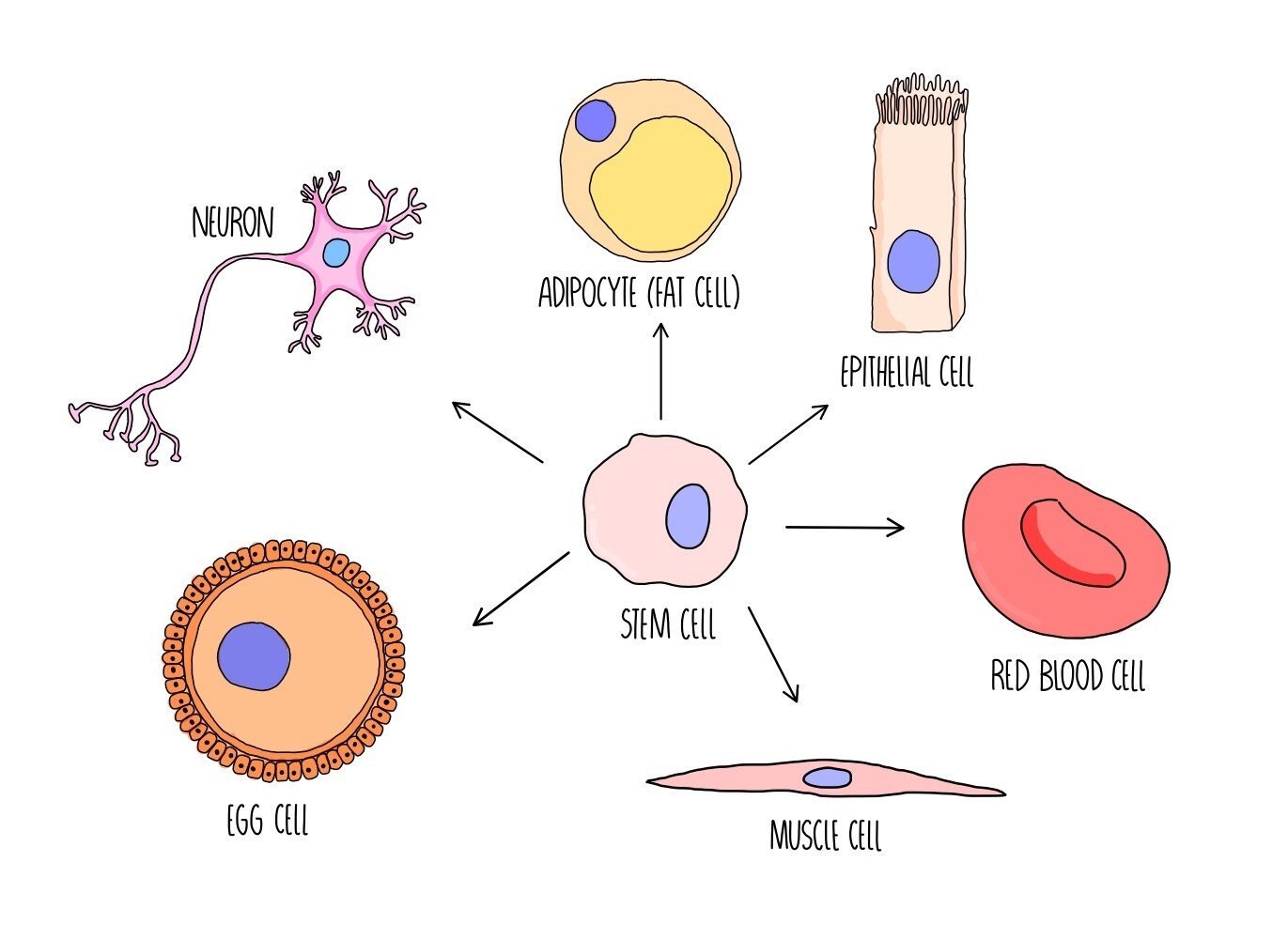
The ability of stem cells to undergo differentiation is referred to as potency - there are different levels of potency:
Totipotent - totipotent cells have the ability to divide into any type of cell (including the extraembryonic cells which make up the placenta and umbilical cord).
Pluripotent - pluripotent cells can divide into any type of cell except the extraembryonic cells.
Multipotent - these cells can divide into a handful of different cell types
Unipotent - these cells can only divide into one type of cell
Gene Expression
All cells of our body contain exactly the same set of genes but can have very different structures and functions in our body. The reason that a brain cell is so much different from a muscle cell is due to the fact that different proteins are being made - this results from the activation (and deactivation) of different genes in these two cells.
Within each cell, certain genes will be activated and others will be inactivated. Only the activated genes are transcribed into mRNA which is translated into protein. The particular proteins that are formed will modify the cell by changing its structure and controlling cellular processes. These changes cause the cell to become specialised.
For example, during the differentiation of a stem cell into a red blood cell, certain genes are activated. These genes are responsible for the production of haemoglobin and of proteins which will destroy the nucleus (enabling the cell to pack in more haemoglobin). Other genes will be inactivated so that any proteins which are unrelated to the functioning of red blood cells will not be produced. The production of red blood cell-related proteins will modify the cell, resulting in a specialised red blood cell.
Transcription Factors
The activation and deactivation of genes is carried out by proteins called transcription factors (TFs). TFs which activate genes are called activators whereas TFs which deactivate genes are referred to as repressors. Activators can work by binding to the beginning of the gene (the promoter region) and helping RNA polymerase to bind and transcribe the gene. Repressors can work by binding to the gene and blocking RNA polymerase from binding.
Operons
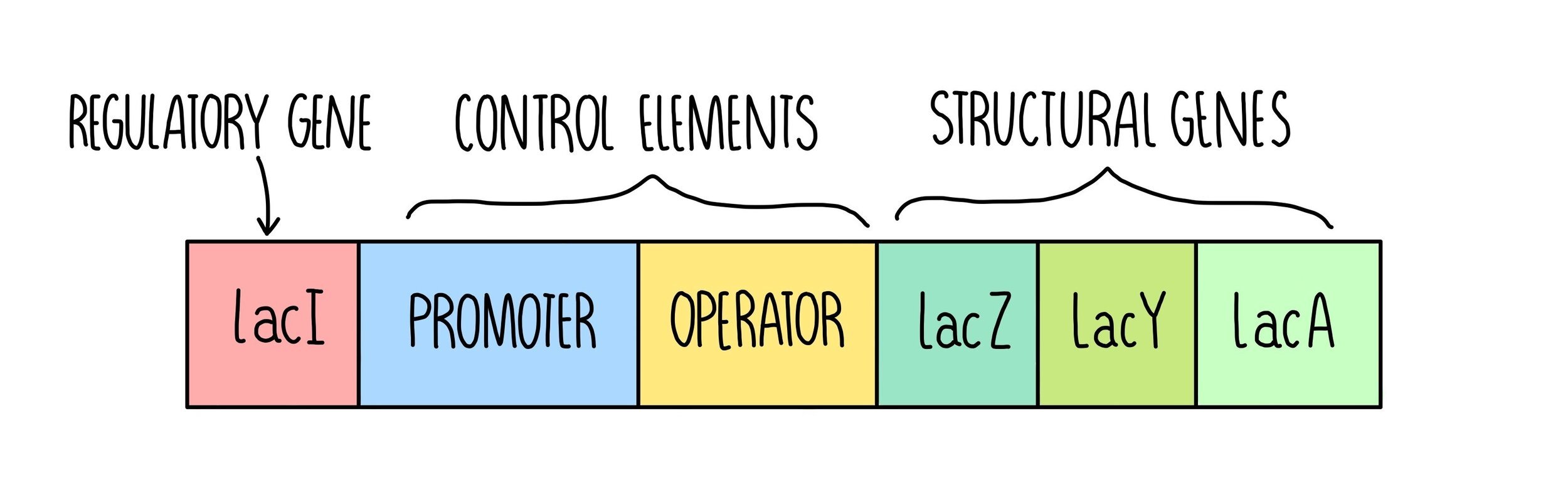
In prokaryotes, transcription factors bind to regions of DNA called operons. An operon is a section of DNA that contains a cluster of genes which are controlled by a single promoter (regulatory region). Operons contain the following elements:
Structural genes - these code for useful proteins such as enzymes
Control elements - these contain a promoter region where RNA polymerase can bind and an operator region where transcription factors can bind
Regulatory gene - these codes for transcription factors (activators or repressors).
Lac Operon
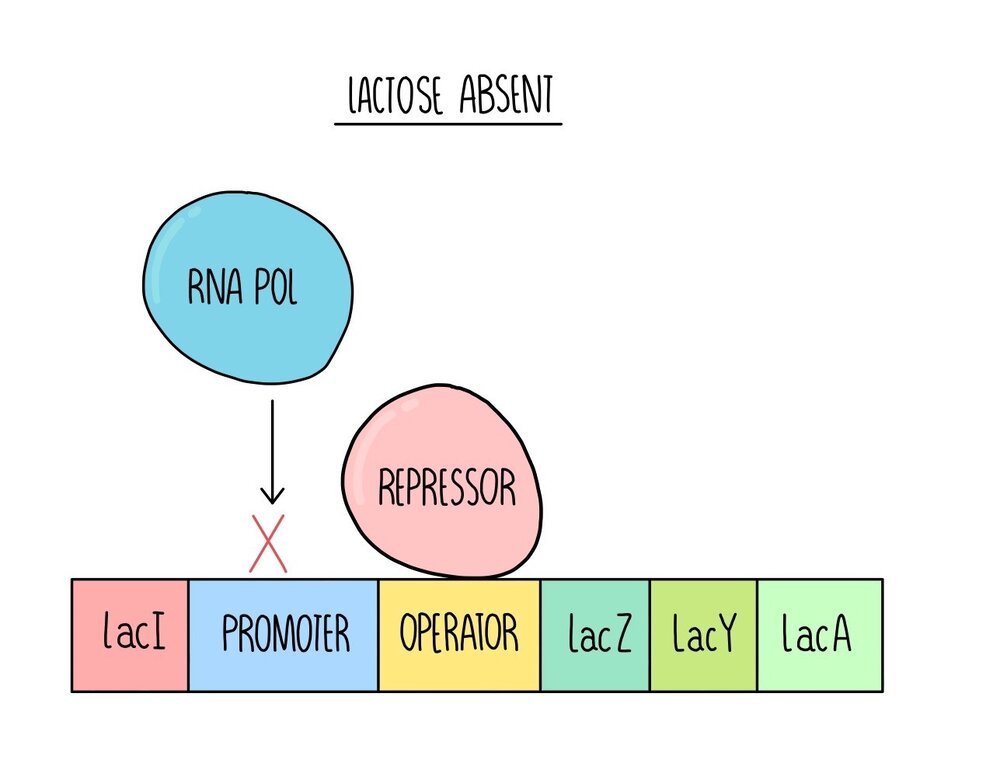
E. coli are a species of bacteria which feed on glucose. When glucose is not available, they can digest lactose. E. coli only produce the enzymes to digest lactose when glucose is absent (and lactose is present), so that they don’t waste energy and resources building proteins that they don’t need. The genes which produce the enzymes to respire lactose are found on an operon called the lac operon.
When lactose is absent, a regulatory gene (lacI) produces a protein called the lac repressor. The lac repressor is a transcription factor which binds to the operator region. This blocks RNA polymerase from binding to the promoter region so the structural genes are not transcribed.

When lactose is present, lactose binds to the repressor and changes its shape. This shape change means that the repressor can no longer bind to the operator, allowing RNA polymerase to bind to the promoter region. RNA polymerase transcribes the structural genes - lacZ, lacY and lacA. These genes code for proteins which allow E. coli to respire lactose. LacZ codes for an enzyme called beta-galactosidase which breaks the glycosidic bond in lactose, breaking it down into glucose and galactose. Lac Y codes for a protein called lactose permease, a membrane proteins which transports lactose into the cell. Scientists still aren’t sure what the function of the lacA gene is and its exact role in digesting lactose.
Stem cells in medicine
Stem cells are being used as a treatment for certain diseases:
Stem cell transplants are given to patients with leukaemia - leukaemia is a type of cancer which destroys stem cells so bone marrow transplants are used to replace the lost stem cells.
Research is being carried out to develop ways of growing whole organs from stem cells. The organs can then be transplanted into the patient to replace organs that have been damaged or are diseased e.g. pancreatic transplants can be given to people with diabetes. This approach will help those who currently have to wait years for an organ donation.
Adult stem cells vs embryonic stem cells
There are two sources of stem cells: adult stem cells (ASCs) and embryonic stem cells (ESCs). ASCs are found in the bone marrowof adults and have a more limited potency. They can only develop into a limited number of cell types (i.e they are multipotent). ESCs are found in human embryos and can develop into all types of adult cell (i.e. they are pluripotent). The ability of ESCs to divide into any cell type makes them much more useful for medicine but the use of ESCs comes with ethical implications, since they are obtained from embryos that have developed in the lab from in vitro fertilisation. ESCs are taken from the embryos when they are 4-5 days old and are then discarded. Some people are opposed to this since they believe that a human has a right to life from the moment of conception.
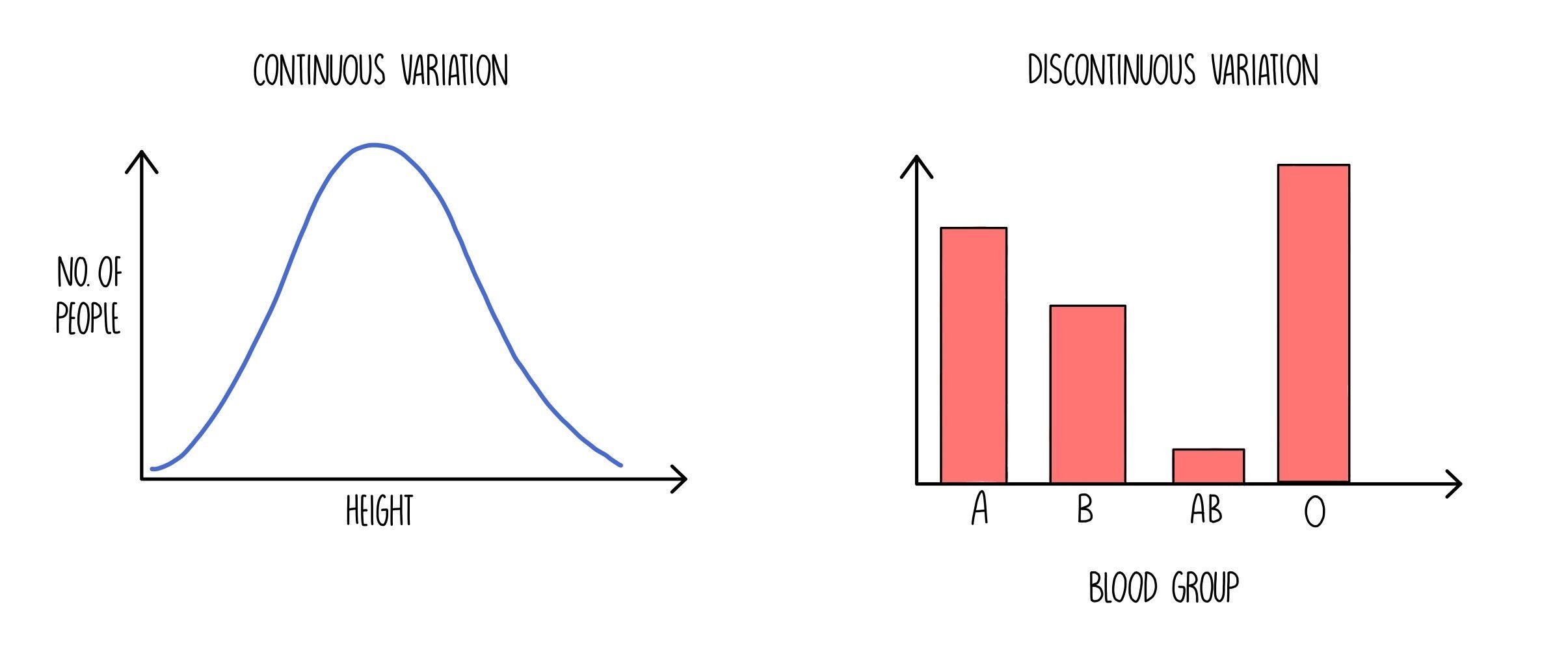
Continuous and discontinuous variation
Characteristics which are controlled by the expression of a single gene are said to be monogenic. These kinds of characteristics typically give rise to discontinuous variation in the phenotype that is expressed - i.e. the phenotypes can be grouped into distinct categories. An example of a characteristic controlled by a single gene is blood type.
However, the majority of characteristics are controlled by many genes at different loci - they are polygenic. Polygenic characteristics show continuous variation which is when the individuals in a population vary within a range. Examples of characteristics which show polygenic inheritance are height, weight and skin colour.
Variation in phenotype is influenced by variations in genotype and the interaction of the environment. Environmental factors can affect both monogenic and polygenic characteristics. For example:
Monoamine oxidase catalyses the breakdown of a chemical called monoamines. A certain allele of a single gene (i.e. monogenic) can cause some people to have lower levels of the monoamine oxidase enzyme, which has been associated with mental health problems. But environmental factors like smoking can also reduce enzyme levels.
Fur colour is a polygenic characteristic often influenced by the environment. Arctic mammals often have dark fur in the summer, which becomes white in the winter for better camouflage. The drop in temperature over the winter changes the expression of certain genes which code for fur pigmentation.
Epigenetics
Epigenetic modification involves the addition or removal of chemical ‘tags’ onto DNA or histone proteins. These modifications are caused by environmental factors (such as diet, stress and smoking) and can occur from as early as when we are in the womb.
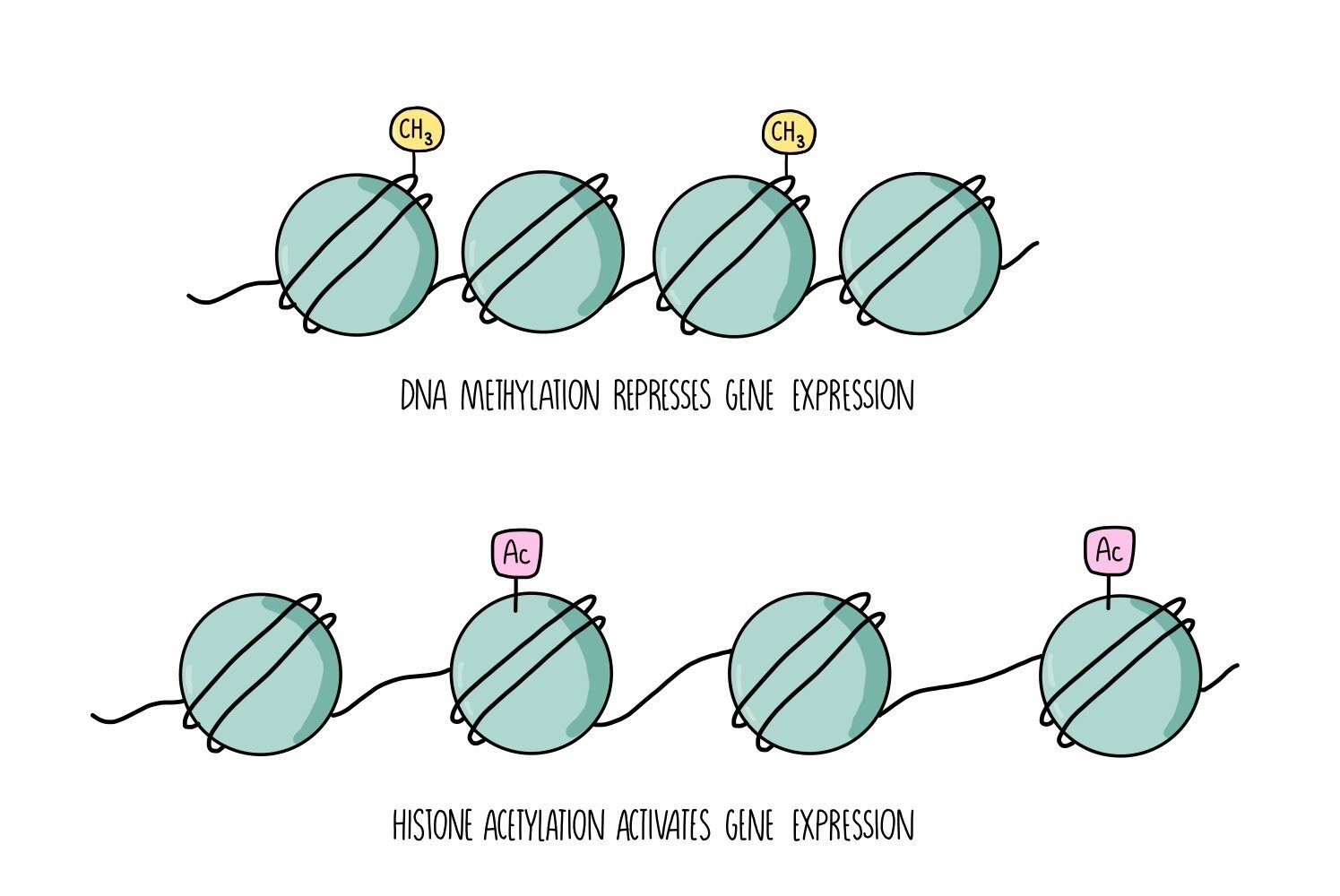
The addition or removal of epigenetic tags changes the structure of the chromosome, making it either more or less accessible to RNA polymerase, transcription factors and other proteins which are involved in transcription. The more open the structure of the chromosome, the more accessible it is to RNA polymerase - the gene is switched on. The less open the structure, the less accessible it is and transcription is reduced - the gene is switched off. The two main types of epigenetic ‘tags’ are methyl groups and acetyl groups.
DNA methylation
Methyl groups (-CH3) can be added directly onto the DNA. They attach to a region of DNA called the CpG site - this is where a cytosine is found next to a guanine. The addition of a methyl group makes the DNA less accessible to the proteins involved in transcription. The gene is switched off (inactivated).
Histone acetylation
Chromosomes are made up of a substance called chromatin, which consists of DNA wrapped around histone proteins. Chromatin can either be tightly wound (condensed) or more loosely wound (less condensed). The more condensed the chromatin, the less accessible it is to the proteins involved in transcription. The addition of acetyl groups to histones causes the chromatin to become less condensed, making it more accessible and activating the gene. The removal of acetyl groups converts the chromatin to a more highly condensed form, which is less accessible and represses transcription of the gene.