Nervous and Hormonal Communication
How the nervous system works
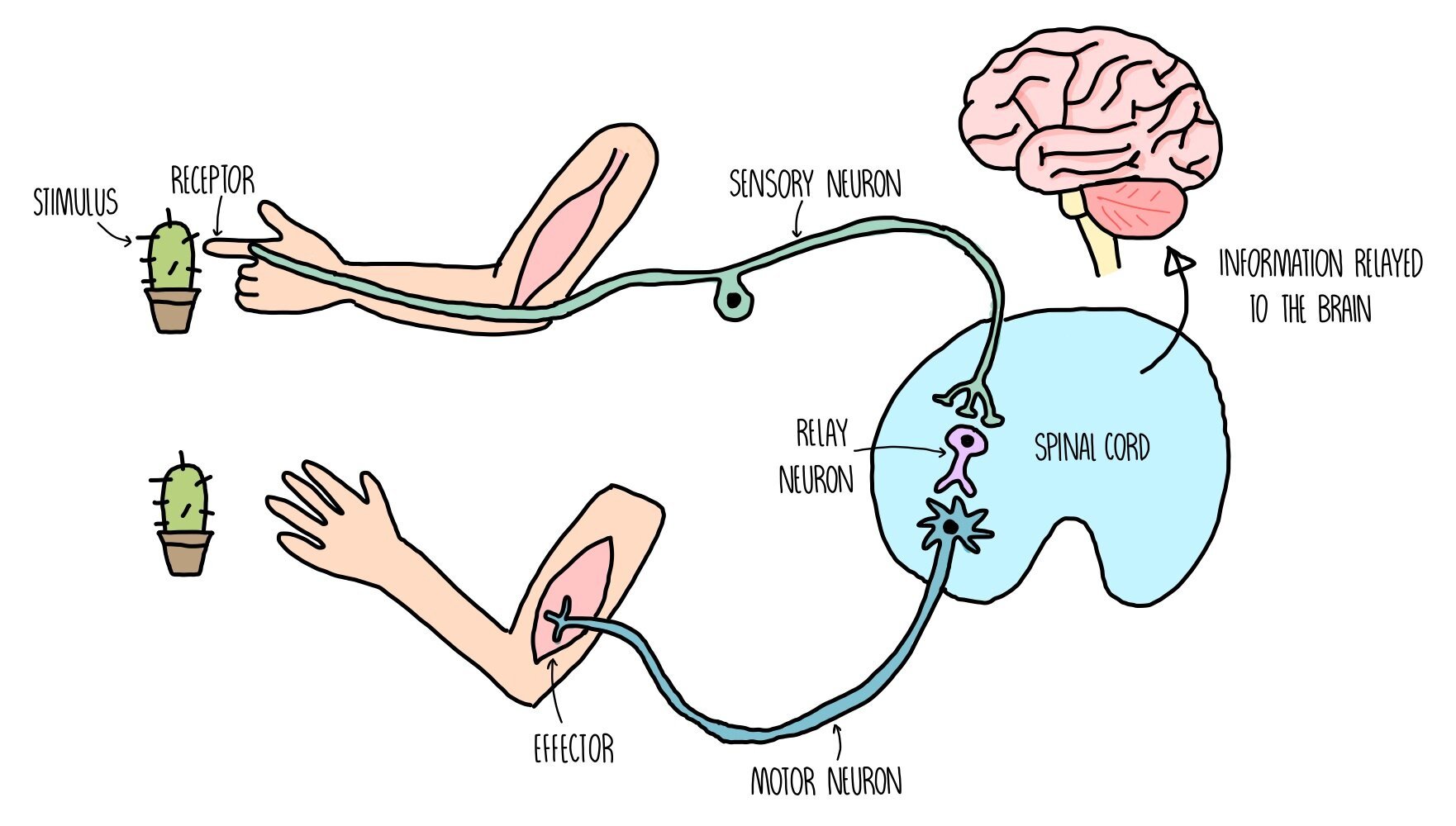
The nervous system detects changes in our environment (known as stimuli) through cells called receptors.
Receptors are sensitive to a number of different aspects of our environment, such as light, pressure (touch) and chemicals in the air (smell).
When receptors detect certain stimuli, they signal to the central nervous system (CNS) through initiating an electrical impulse through a neuron (nerve cell).
The neuron which sends an electrical impulse from the receptor within the sense organ and the coordination centre is called the sensory neuron.
The coordination centre receives impulses from various receptors around the body, processes the information and coordinates a response by signalling to other parts of the body. Coordination centres include the brain, spinal cord and pancreas.
These organs will signal to an effector (a muscle or gland) by releasing an electrical impulse along a motor neuron.
Stimulation of an effector will produce a response such as muscle contraction or hormonal release.
How the hormonal system works
The endocrine (hormonal) system is a group of glands, which secrete chemicals called hormones.
Glands secrete hormones when they are stimulated by neurons or by a change in concentration of a chemical substance (such as another hormone).
Hormones travel around our body in our bloodstream to their target organs where it produces an effect.
Target cells possess complementary receptors on their cell surface membranes that allow it to respond to the hormone, triggering a response inside the cell.
Hormones can be proteins/peptides e.g. insulin or steroids (a type of lipid) e.g. testosterone.
Compared to the nervous system, the endocrine system is:
o Slower to bring about an effect
o Longer-lasting
o Widespread response (can act on multiple target tissues)
o Involves the use of chemicals rather than electrical impulses
Receptors
Our nervous system uses receptors to detect stimuli (changes in the environment) and pass on this information to the CNS. Receptors can either be whole cells (e.g. photoreceptors are cells which are sensitive to light) or proteins molecules which are found on the cell surface membrane. Each receptor is specific to a single type of stimulus, such as light, temperature or glucose concentration. When a receptor is not stimulated, there is a charge difference between the inside and outside of the membrane and it is said to be polarised. When the receptor detects a stimulus, the permeability of its cell membrane changes which changes the charge difference (potential difference) across the membrane. If the change in potential difference is large enough (i.e. it exceeds the threshold level), it will trigger an action potential (an electrical impulse) in a sensory neuron.
We contain the following receptors in our sense organs:
Chemoreceptors - receptors which detect chemicals
Thermoreceptors - receptors which detect heat
Mechanoreceptors - receptors which detect pressure (e.g. Pacinian corpuscles)
Photoreceptors - receptors which detect light (e.g. rods and cones)
Photoreceptors
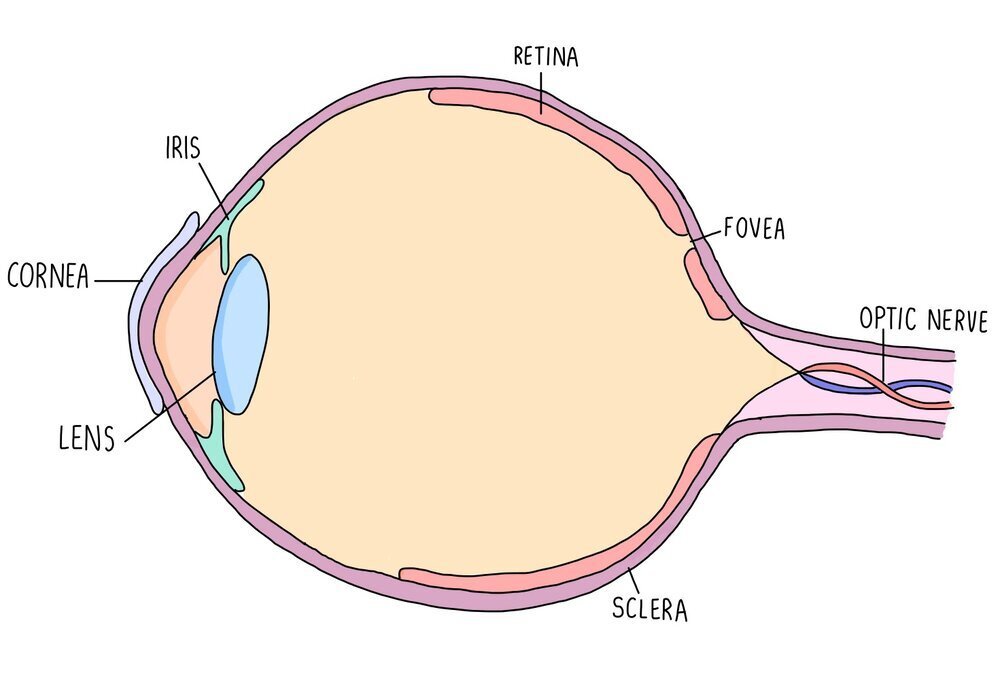
Photoreceptors are receptors which detect light and are found in the retina of the eye. There is an area of the retina called the fovea which contains a cluster of photoreceptor cells. Photoreceptors detect light as it hits the retina and send nerve impulses to the brain along the optic nerve. The region of the eye containing the optic nerve is called the blind spot since there are no photoreceptors in this region so light can’t be detected. Photoreceptors are connected to the optic nerve through a bipolar neurone.
The human eye has two types of photoreceptors - rods and cones. Rods are mostly located along the outside of the retina while cones are clustered together in the fovea. Rods are responsible for black-and-white vision and can function in lower light levels than cones. They are much more sensitive than cones, so are the type of photoreceptor used for visualising objects in the dark. Cones are responsible for colour vision and are sensitive to blue, green or red light. Different cones are stimulated in different proportions, so that we see different colours. Cone cells provide good visual acuity (the ability to distinguish between two points which are close together) because each cone cell has its own synapse via a bipolar neurone which connects to the optic nerve.

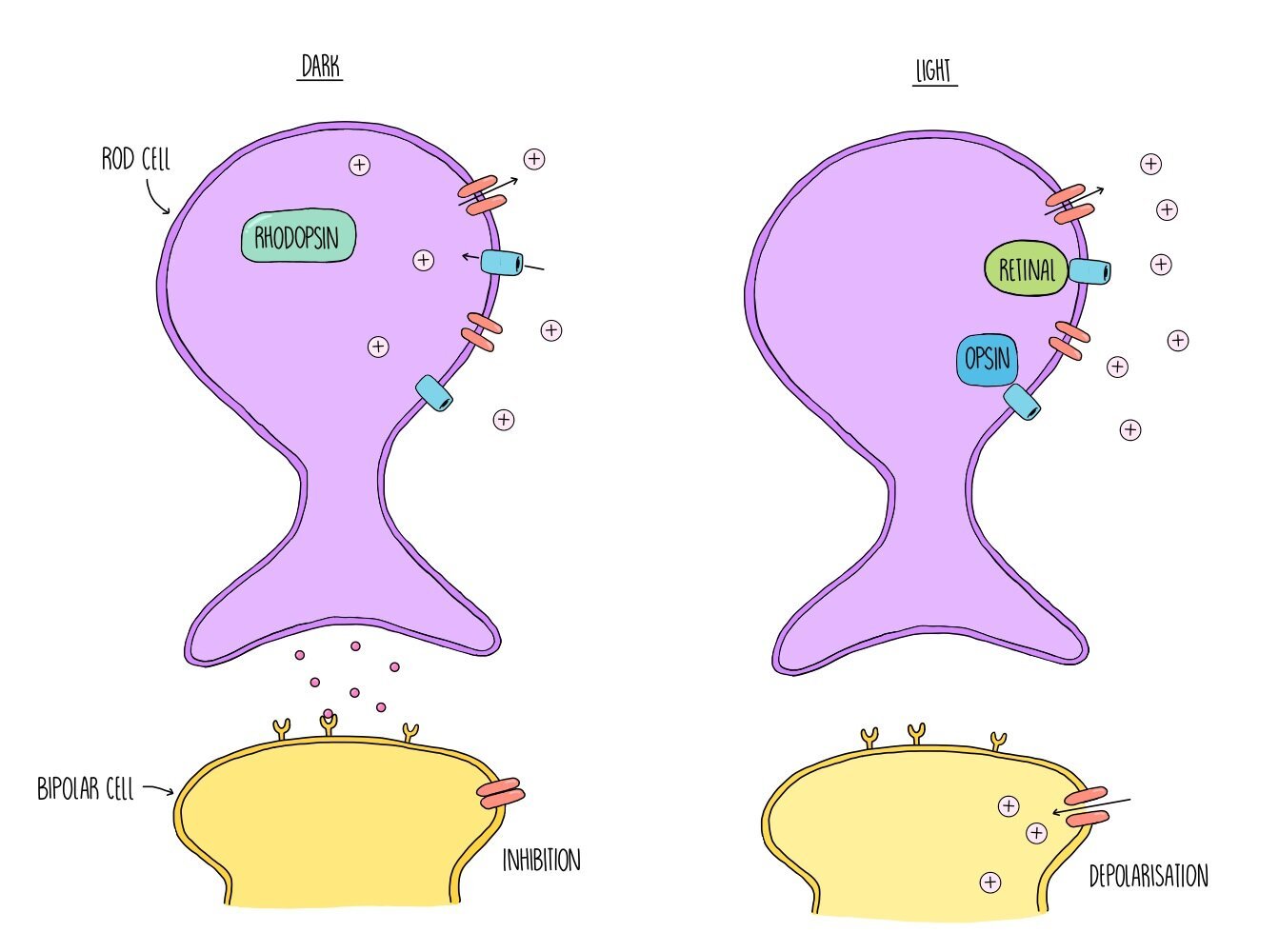
In dark conditions, the membrane of rod cells is depolarised, which means there is not much difference in charge between the inside and outside of the membrane. This is because the rod cells actively transport sodium ions out of the cell, which flow straight back into the cell through sodium ion channels. Depolarisation of the rod cell membrane triggers the release of neurotransmitters which inhibit the bipolar neurone. The bipolar neurone cannot fire an action potential which means that no information is sent to the brain. However, when light is present, light energy causes a pigment called rhodopsin to split apart into two proteins, retinal and opsin. This process is referred to as the bleaching of rhodopsin and it causes sodium ion channels in the cell surface membrane to close. Sodium ions continue to be actively transported out of the rod cells but they cannot flow back into the cell through the ion channels, creating a difference in charge across the membrane. The inside of the membrane is now much more negative compared to the outside and the rod cell is said to be hyperpolarised. When the rod cell is hyperpolarised it stops releasing neurotransmitters. This means that inhibition of the bipolar neurone stops and it can now become depolarised. If depolarisation exceeds a threshold level, information is passed onto the brain via the optic nerve and the presence of light is detected.
Types of neuron
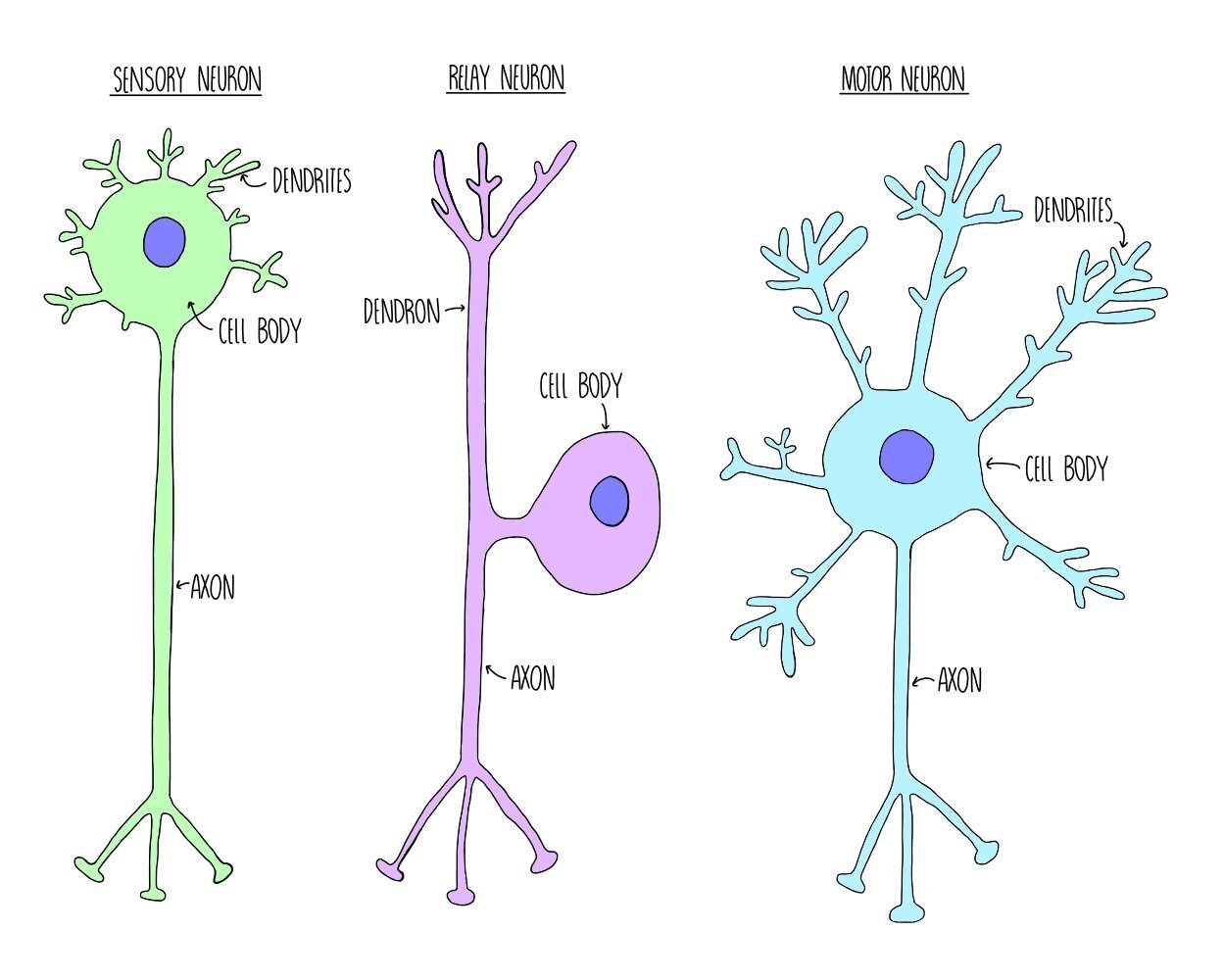
Neurons are cells which carry information to and from the central nervous system, in the form of electrical impulses called action potentials. There are three different types of neurone, with slightly different structures. What they all have in common, however, is a cell body containing a nucleus, dendrites which carry an action potential towards the cell body and an axon which carries the action potential away from the cell body.
Sensory neurons carry action potentials from receptors to the central nervous system. They consist of one long dendron and a short axon.
Relay neurons carry action potentials between the sensory and motor neurons and are found within the CNS. They have lots of short dendrites.
Motor neurons carry action potentials from the CNS to an effector. They have lots of short dendrites and one long axon.
Resting potential
When a neurone is not firing (i.e. it is not transmitting an action potential), there is a difference in charge between the inside and the outside of the membrane (it is polarised). This charge difference is referred to as the resting potential and is usually around -70 mV. Polarisation of neuronal cell membranes at rest occurs due to the action of sodium-potassium ion pumps. These pumps are found within the cell membrane and actively transport sodium and potassium ions into and out of the neurone. For every three sodium ions that the proteins pump out of the cell, they pump two potassium ions into the cell. This ensures that there are always more positive ions out of the cell compared to inside the cell and makes sure there is a charge difference across the membrane.

Action potential
When a neurone is stimulated, the charge difference between the inside and outside of the cell membrane is lost and the membrane is depolarised. If enough charge is lost and depolarisation exceeds -55 mV, an action potential will occur in that neurone. The -55 mV ‘limit’ is known as the threshold potential - any depolarisation above this number will result in an action potential whereas anything less than that will result in nothing. We therefore refer to action potentials as an “all-or-nothing” response.
Depolarisation during an action potential occurs because sodium ion channels open up in the membrane. Remember that the sodium-potassium ion pump has been actively transporting sodium ions out of the neurone, creating a sodium ion concentration gradient. This means that when sodium ion channels open, sodium ions flood into the neuron by facilitated diffusion. The potential difference across the membrane is reduced until it reaches a voltage of around +30 mV.
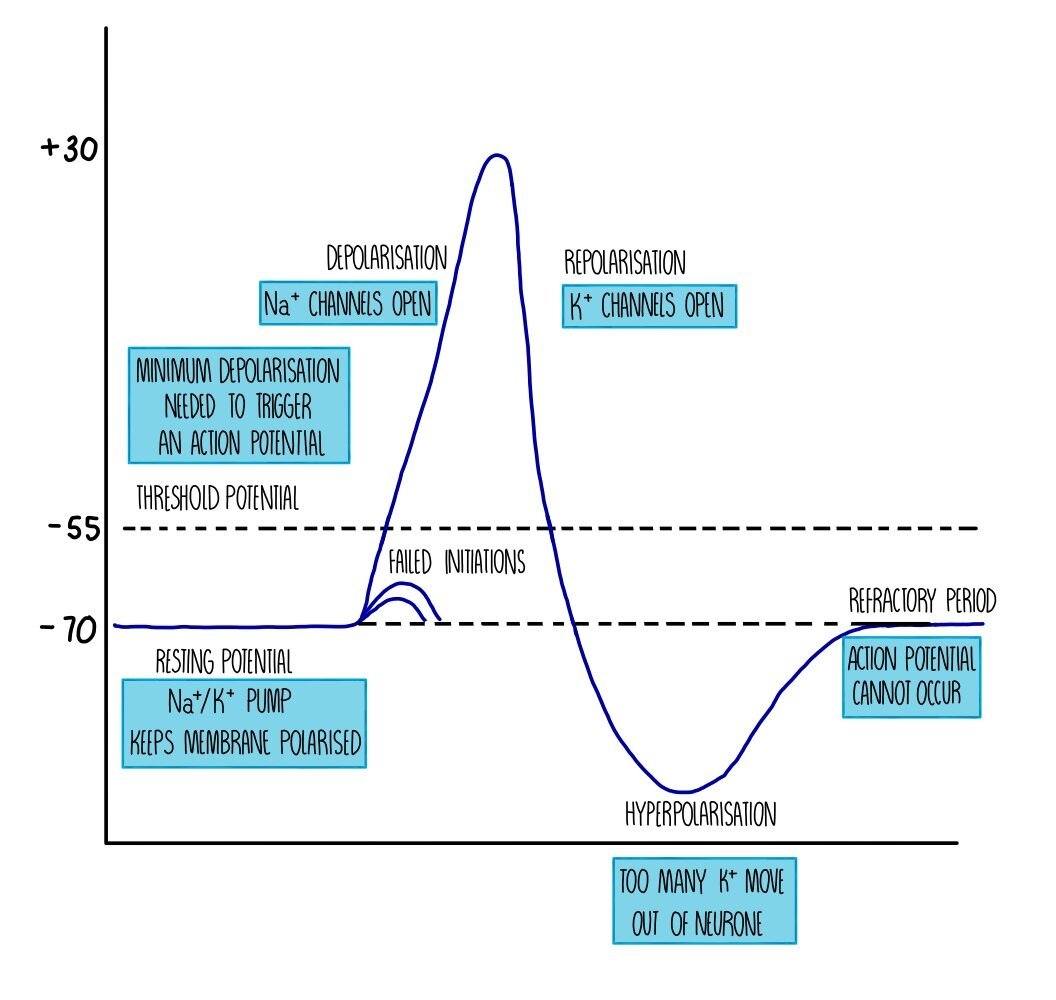
Sodium ion channels close and potassium ion channels open, which causes potassium ions to move out of the neurone down their concentration gradient. The movement of positive ions out of the cell means that there is a charge difference again across the membrane - this is called repolarisation. However, the charge difference exceeds the resting potential and becomes ‘hyperpolarised’. This is because the potassium ion channels are slow to close and too many potassium ions diffuse out of the neurone. The action of the sodium-potassium ion pump restores the balance between sodium and potassium ions on either side of the membrane and returns the neurone to its resting potential of -70 mV.
Immediately after an action potential is a brief period called the refractory period. During the refractory period, the neurone cannot be stimulated and an action potential cannot occur. This is because the ion channels are recovering and they cannot be made to open. The refractory period is important because it ensures that action potentials do not overlap (i.e. they pass along the neurone as separate impulses) and that action potentials are unidirectional.
Once an action potential occurs in one part of the neuron, it will stimulate an action potential in the adjacent part of the neuron, creating a kind of ‘Mexican wave’ of depolarisation. This wave of depolarisation occurs because the sodium ions which diffuse into the neuron diffuse sideways, causing voltage-gated ion channels in the next portion of the neurone to open, so sodium ions move into the neurone further along the membrane. The wave moves away from the part of the neurone which has just fired an action potential because that part of the neurone will be in the refractory period and cannot be stimulated.
Saltatory conduction
Some neurones are insulated with a fatty layer along the axon. This fatty layer is called a myelin sheath and is made up of a type of cell called a Schwann cell. The myelin sheath acts as an electrical insulator, which means that ions cannot move into or out of the myelinated portions of the neurone. However, there are gaps in the myelin sheath called nodes of Ranvier, where sodium ion channels and potassium ion channels are concentrated. Action potentials occur only at the nodes of Ranvier - when one node is stimulated this triggers depolarisation of the next node, causing the impulse to ‘jump’ from node to node. This type of nervous transmission is called saltatory conduction and is much faster than transmission along non-myelinated neurones, where the action potential has to travel along the entire length of the neurone in a wave of depolarisation. The speed at which an action potential moves along a neurone is known as the conduction velocity - the higher the conduction velocity, the faster the action potential is travelling. This means that action potentials along myelinated neurones have a higher conduction velocity compared to those travelling along non-myelinated neurones.
Size of the stimulus
An action potential is an ‘all-or-nothing’ response. If the threshold potential is reached, an action potential will occur. This action potential is always of the same voltage (depolarisation to +30 mV) regardless of whether the stimulus that initiated the action potential is small (e.g. a pinprick) or large (e.g. a sledgehammer). If the threshold isn’t reached then an action potential will not be fired. The difference between action potentials resulting from stimuli of different sizes is the frequency that action potentials are firing - the bigger the stimulus, the more often an action potential will occur along the neurone.
Anaesthetics
Anaesthetics are drugs which create a numbing sensation and are used in medicine to prevent patients from feeling pain (e.g. during an operation). They work by binding to sodium ion channels in the neurone and preventing them from opening. If sodium ions cannot move into the neurone, then the membrane cannot depolarise and an action potential cannot occur. This prevents neurone from sending a pain impulse to the brain, so the brain doesn’t register anything.
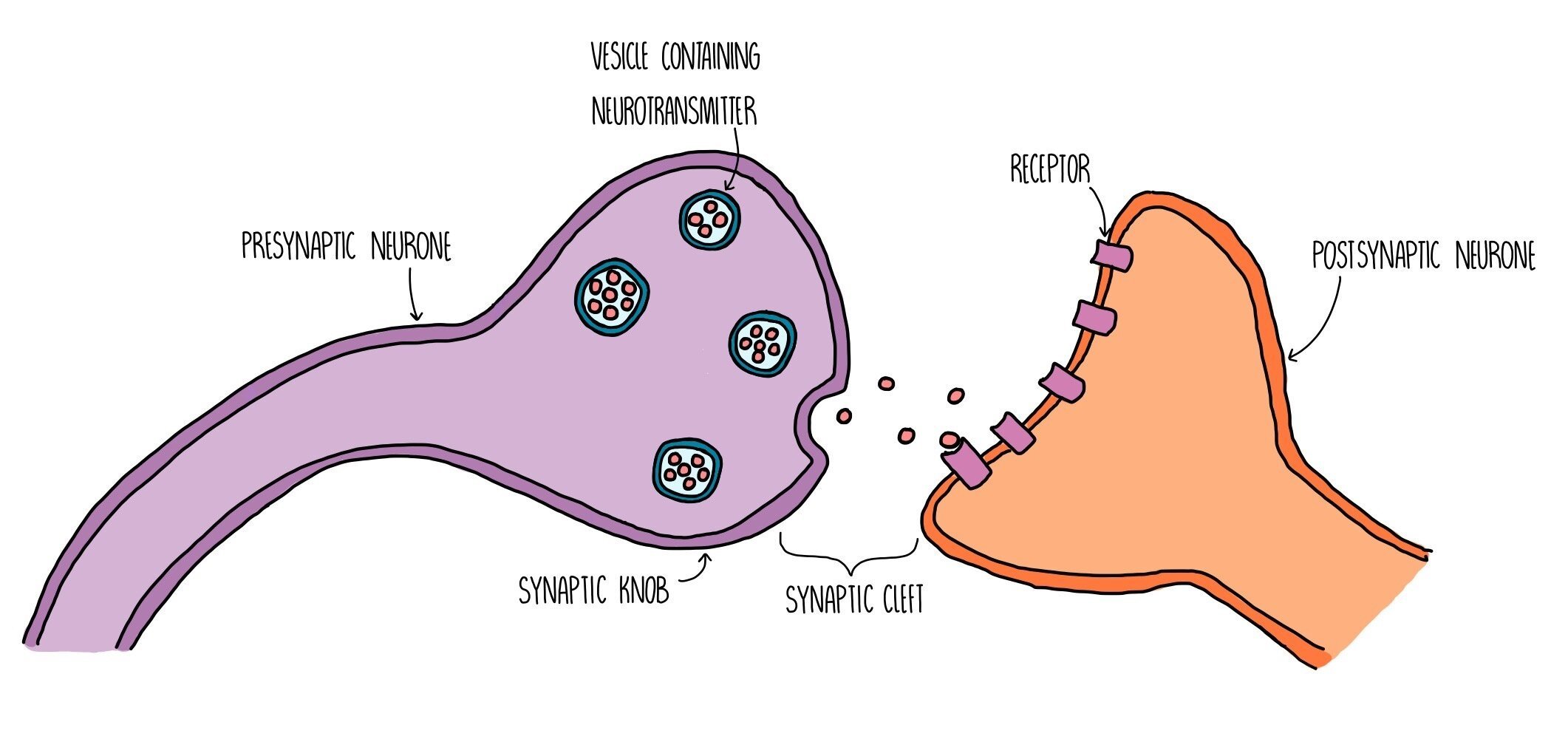
Synapses
A synapse is a gap found between neurones (or between a motor neurone and an effector). Electrical impulses cannot pass through the gap, so neurones release neurotransmitters from one neurone to the next to stimulate an action potential in the next neurone. The neurone before the synapse is called the presynaptic neurone and the one after the synapse is called the postsynaptic neurone. The space between them is called the synaptic cleft. This means that action potentials will travel along the presynaptic neurone, through the synaptic cleft (via neurotransmitters) then along the postsynaptic neurone. The presynaptic neurone has a swelling at the end which is called the synaptic knob.
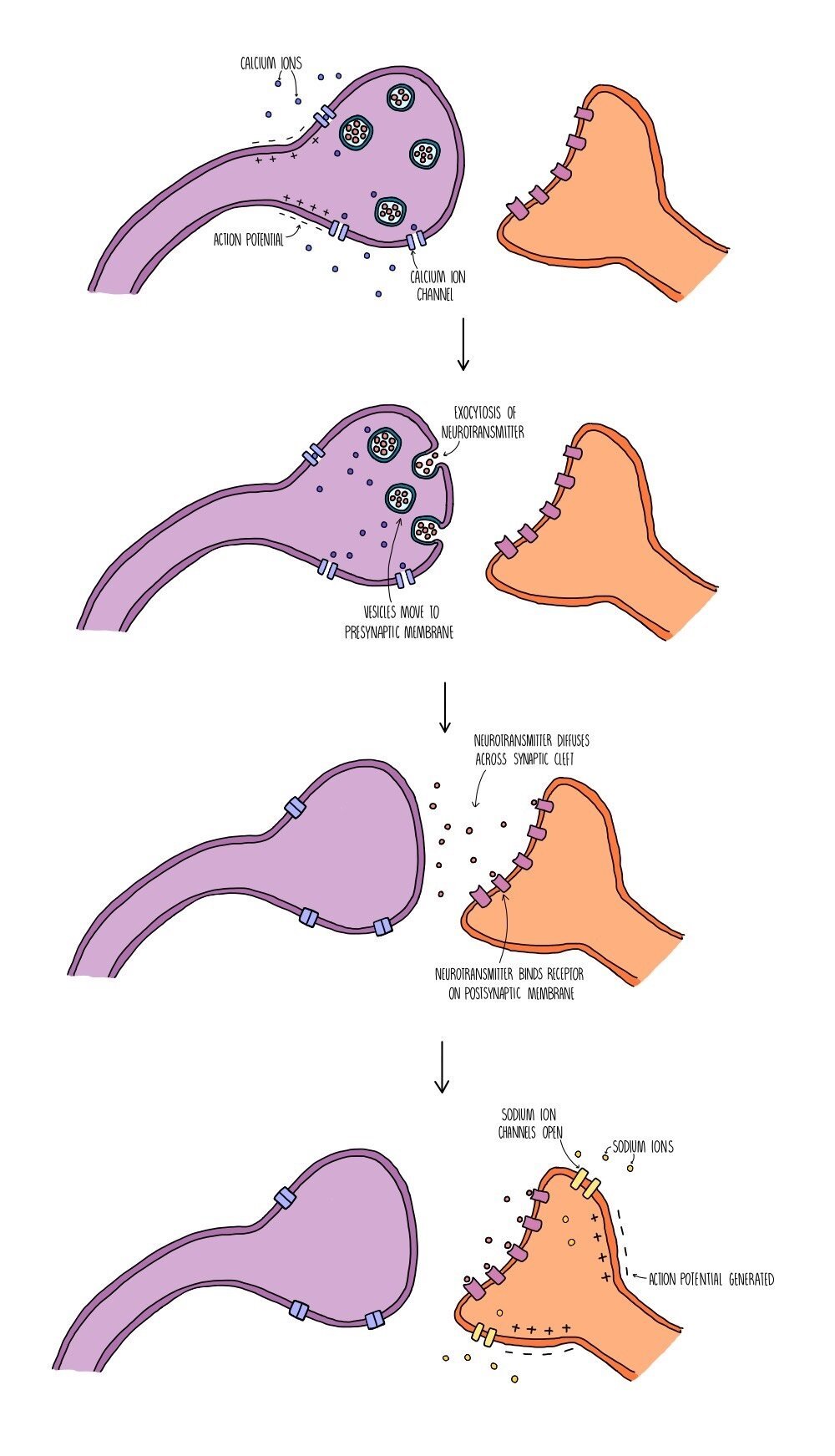
Synaptic transmission takes place in the following stages:
An action potential arrives at the end of the presynaptic neurone (at the synaptic knob) and triggers the opening of voltage-gated calcium ion channels.
Calcium ions move into the synaptic knob by facilitated diffusion and trigger the movement of vesicles containing neurotransmitters (such as acetylcholine or dopamine) towards the presynaptic membrane.
The vesicles fuse with the presynaptic membrane and their contents is released by exocytosis.
The neurotransmitters diffuse across the synaptic cleft and bind to specific receptors on the postsynaptic membrane.
This triggers the opening of sodium ion channels in the postsynaptic membrane. Sodium ions move into the postsynaptic neurone, causing depolarisation and triggering an action potential if the excitation exceeds the threshold potential of -55 mV.
The neurotransmitter is removed from the synaptic cleft which prevents the continuous stimulation of an action potential in the postsynaptic neurone. The neurotransmitter is either reabsorbed by the presynaptic neurone (and recycled) or broken down by enzymes in the synaptic cleft (and the products are reabsorbed).
Receptors which are complementary to neurotransmitters, such as acetylcholine, are only found on the postsynaptic membrane - there are none within the postsynaptic membrane). This ensures that the action potential always travels in one direction only (unidirectional) and prevents the nerve impulse from travelling backwards.
Synaptic divergence and convergence
When one neurone forms connections to multiple neurones through multiple synapses, the action potential can diverge, sending information to different parts of the body. This is known as synaptic divergence. On the other hand, multiple neurones can all connect to a single neurone, causing the action potentials from multiple neurones to converge and become amplified (synaptic convergence). This creates a stronger impulse when many different neurones are activated that form part of the same neural pathway.

Tropisms
Tropisms are responses by an organism in a particular direction as a result of an external stimulus. Plants show phototropism (growth responses to sunlight) and geotropism (growth responses to the force of gravity). These tropisms are a result of growth factors which accumulate in different parts of the plant.
Phototropism is a directional response to sunlight. Plant shoots show positive phototropism and grow towards the sun. This maximises the amount of light they can absorb for photosynthesis. Roots show negative phototropism and grow away from the sun. This ensures that the roots bury themselves deeper within the soil, so they can absorb more water for photosynthesis.
Geotropism is a directional response to gravity. Plant shoots show negative geotropism and grow away from the force of gravity. This ensures that plants grow upwards and means that more light is absorbed for photosynthesis. Roots show positive geotropism and grow towards the force of gravity. Again, this ensures that roots bury themselves deeper within the soil, so they can anchor the plant and absorb more water for photosynthesis.
Growth factors
Plants produce different growth factors which enable them to respond to their environment. Growth factors are produced in regions of the plant that are actively dividing (i.e. the shoots and leaves) and diffuse to other parts of the plant where they are needed. Auxins promote cell elongation in shoots but have the opposite effect in roots (auxins accumulate within roots in high concentrations which inhibits growth).
One of the most important groups of growth factors are auxins such as indoleacetic acid, which is involved in both phototropism and geotropism. There are other growth factors which also play important roles in plant growth:
Gibberellins stimulate seed germination and flowering.
Abscisic acid (ABA) helps plants respond to environmental stress and is involved in stomatal closure.
Cytokinins stimulate cell division and cell differentiation.
Ethene stimulates flowering and fruit ripening.
Indoleacetic acid (IAA)

Indoleacetic acid (IAA) is a type of auxin which allows plants to response to light (phototropism) and gravity (geotropism). It works by entering the nucleus of plant cells and binds to the promoter regions of DNA. It then acts as a transcription factor, activating or inhibiting the transcription of genes which code for proteins involved in cell elongation and growth. IAA is transported around the plant in the phloem then moves shorter distances within the plant (from cell to cell) by diffusion and active transport.
In shoots, IAA accumulates on the shaded part of the shoot where it activates genes involved in cell elongation. The activated genes are transcribed into proteins which make the cell walls looser and stretchier, causing the cells to become longer. The cells on the shaded side of the stem are longer than those on the sunny side, causing the shoot to bend towards the sun.
In roots, IAA also accumulates on the more shaded side but this time inhibits the growth of cells. This means more cells are produced on the non-shaded side of the root, causing the root to bend away from the sun.
IAA also regulates geotropism by accumulating on the underside of shoots and roots. In shoots, IAA causes cell elongation on the underside of the shoot, causing it to bend upwards, away from the force of gravity. In roots, IAA inhibits cell growth, causing roots to grow downwards, towards the force of gravity.
Phytochromes

Plants don’t just respond to sunlight and gravity - they can also tell whether it is daytime or night-time and even what season it is using special photoreceptors (receptors which detect light) called phytochromes. Phytochromes are found in the leaves, seeds, roots and stem of plants and exist in one of two states. The PR state absorbs light with a wavelength of 660 nm (the red part of the visible spectrum) and the PFR state absorbs light with a wavelength of 730 nm (the far-red part of the visible spectrum).
When PR absorbs red light, it is quickly converted into the PFR state and when PFR absorbs far-red light, it is quickly converted into the PR state. PFR also turns into PR when there is no light (i.e. during the night).
Because daylight contains more red light than far-red light, more PR is converted into PFR than the other way around. This means that there is more PFR in the daytime and more PR at night. The proportion of PR to PFR allows the plant to 'know' whether it is daytime, night-time, summer or winter. For example, high levels of PFR can stimulate flowering. PFR accumulates during the summer, when there are short nights so less conversion of PFR into PR. High PFR stimulates the transcription of genes involved in flowering, which means that flowering only occurs in the summer.