Responding to Stimuli
Responding to the environment
Organisms respond to their external environment to increase their chances of survival e.g. running away from a hungry lion. They also respond to their internal environment to ensure the conditions are optimal for metabolism (e.g. eating in response to hunger, or shivering when cold). A change in the internal or external environment is called a stimulus.
How the nervous system works
The nervous system detects changes in our environment (known as stimuli) through cells called receptors.
Receptors are sensitive to a number of different aspects of our environment, such as light, pressure (touch) and chemicals in the air (smell).
When receptors detect certain stimuli, they signal to the central nervous system (CNS) through initiating an electrical impulse through a neuron (nerve cell).
The neuron which sends an electrical impulse from the receptor within the sense organ and the coordination centre is called the sensory neuron.
The coordination centre receives impulses from various receptors around the body, processes the information and coordinates a response by signalling to other parts of the body. Coordination centres include the brain, spinal cord and pancreas.
These organs will signal to an effector (a muscle or gland) by releasing an electrical impulse along a motor neuron.
Relay neurons transmit impulses between sensory and motor neurons.
Stimulation of an effector will produce a response such as muscle contraction or hormonal release.
This process occurs when we need to respond immediately to harmful stimuli in our environment, such as accidentally touching a hot object. These unconscious responses are called reflex actions and protect our body from harm through a coordinated response which bypasses the brain. The information is sent directly to the spinal cord, where the electrical impulse is passed from the sensory neuron to motor neuron via a relay neuron. Reflex actions are extremely quick and do not require any conscious input from the brain.

For example, withdrawing your hand from a hot object involves a reflex response:
Thermoreceptors in your hand detect the high temperature (stimulus)
An impulse is sent along a sensory neuron to the spinal cord
A relay neuron transfers the impulse to a motor neuron
A motor neuron delivers the impulse to the arm muscle (effector)
The arm muscle contracts to pull the arm away from the hot object (response)
Tropisms
Tropisms are responses by an organism in a particular direction as a result of an external stimulus. Plants show phototropism (growth responses to sunlight) and geotropism (growth responses to the force of gravity). These tropisms are a result of growth factors which accumulate in different parts of the plant.
Phototropism is a directional response to sunlight. Plant shoots show positive phototropism and grow towards the sun. This maximises the amount of light they can absorb for photosynthesis. Roots show negative phototropism and grow away from the sun. This ensures that the roots bury themselves deeper within the soil, so they can absorb more water for photosynthesis.
Geotropism is a directional response to gravity. Plant shoots show negative geotropism and grow away from the force of gravity. This ensures that plants grow upwards and means that more light is absorbed for photosynthesis. Roots show positive geotropism and grow towards the force of gravity. Again, this ensures that roots bury themselves deeper within the soil, so they can anchor the plant and absorb more water for photosynthesis.
Growth factors
Plants produce different growth factors which enable them to respond to their environment. Growth factors are produced in regions of the plant that are actively dividing (i.e. the shoots and leaves) and diffuse to other parts of the plant where they are needed. Auxins promote cell elongation in shoots but have the opposite effect in roots (auxins accumulate within roots in high concentrations which inhibits growth).
One of the most important groups of growth factors is auxins such as indoleacetic acid, which is involved in both phototropism and geotropism. There are other growth factors which also play important roles in plant growth:
Gibberellins stimulate seed germination and flowering.
Abscisic acid (ABA) helps plants respond to environmental stress and is involved in stomatal closure.
Cytokinins stimulate cell division and cell differentiation.
Ethene stimulates flowering and fruit ripening.
Indoleacetic acid (IAA)
Indoleacetic acid (IAA) is a type of auxin which allows plants to response to light (phototropism) and gravity (geotropism). It works by entering the nucleus of plant cells and binds to the promoter regions of DNA. It then acts as a transcription factor, activating or inhibiting the transcription of genes which code for proteins involved in cell elongation and growth. IAA is transported around the plant in the phloem then moves shorter distances within the plant (from cell to cell) by diffusion and active transport.
In shoots, IAA accumulates on the shaded part of the shoot where it activates genes involved in cell elongation. The activated genes are transcribed into proteins which make the cell walls looser and stretchier, causing the cells to become longer. The cells on the shaded side of the stem are longer than those on the sunny side, causing the shoot to bend towards the sun.
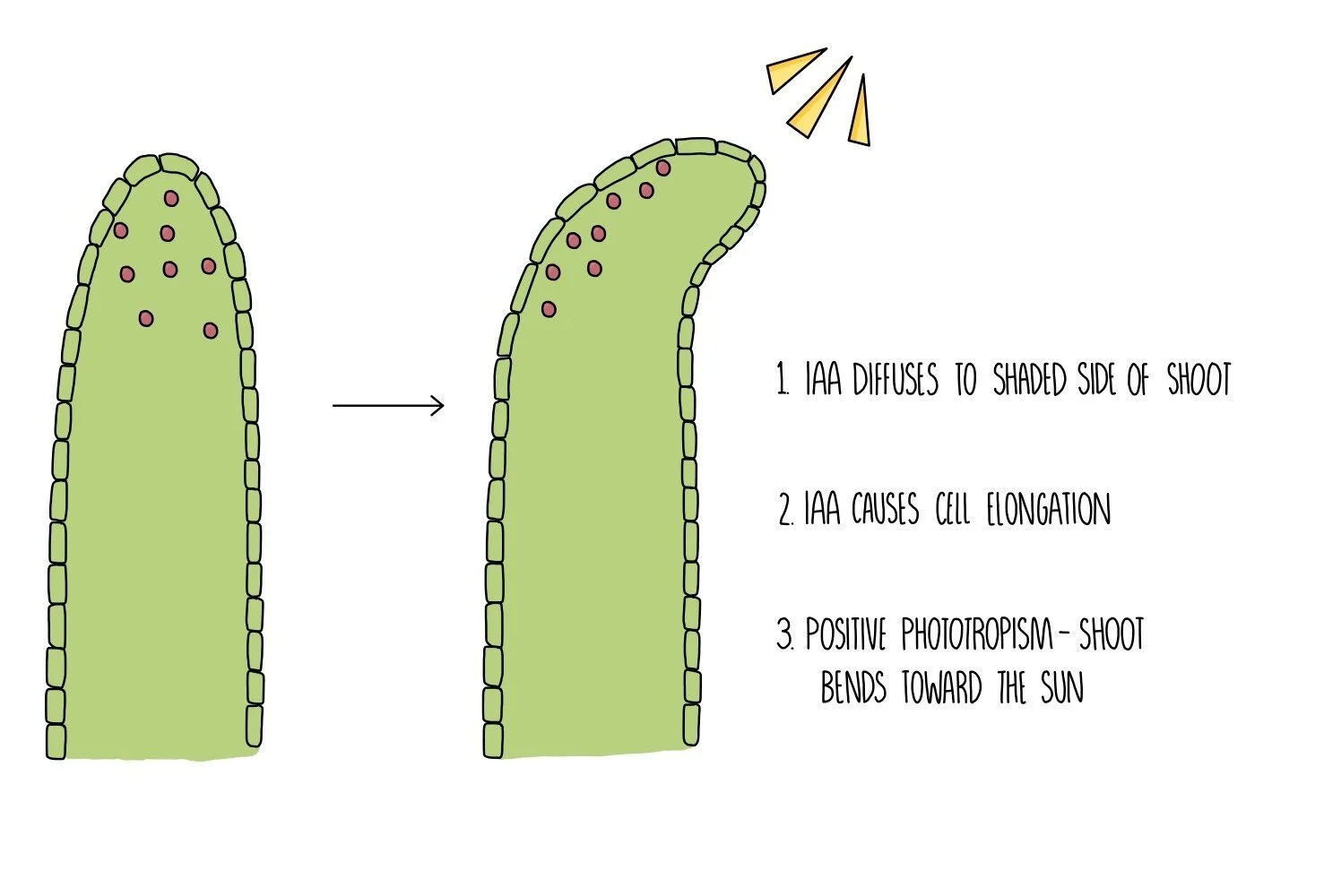
In roots, IAA also accumulates on the more shaded side but this time inhibits the growth of cells. This means more cells are produced on the non-shaded side of the root, causing the root to bend away from the sun.
IAA also regulates geotropism by accumulating on the underside of shoots and roots. In shoots, IAA causes cell elongation on the underside of the shoot, causing it to bend upwards, away from the force of gravity. In roots, IAA inhibits cell growth, causing roots to grow downwards, towards the force of gravity.
Taxes and kineses

Small, mobile organisms such as woodlice exhibit simple responses that optimise environmental conditions – these are grouped as taxes or kineses.
Taxes are where the organism moves towards or away from a directional stimulus. For example, woodlice display negative phototaxis, which means they move away from light. This aids survival as it keeps them hidden under rocks and out of view from predators. It also keeps them in damp conditions and reduces the risk of dehydration.
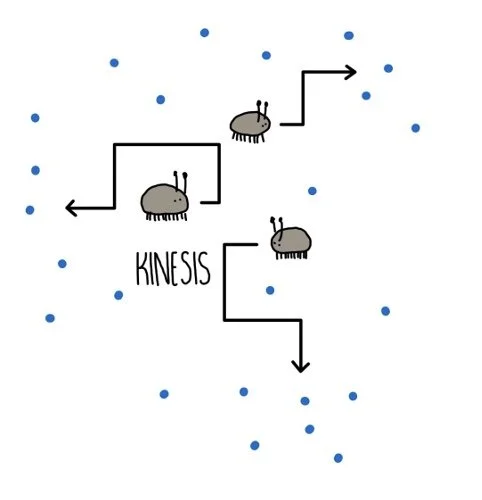
Kineses are movements in response to a non-directional stimulus. For example, woodlice display a kinetic response to humidity. When humidity is high, woodlice move around slowly and don’t change direction much. But when humidity drops, their pace speeds up and they change direction more frequently. This increases the chances that they will find themselves somewhere with more moisture in the air, increasing their chances of survival.
Investigating animal responses using choice chambers
A choice chamber is an apparatus containing different environmental conditions in different compartments. It can be used to investigate how animals respond to light intensity or humidity in the lab.
To investigate the effect of light intensity on woodlouse movement:
Cover one half of the choice chamber, including the sides, with black paper. Line the base with damp filter paper.
Place ten woodlice on the mesh in the centre of the chamber and cover with a lid.
Wait ten minutes, then take off the lid and count the number of woodlice on each side of the chamber.
Carefully move the woodlice back to the centre and repeat the experiment.
The woodlice should display a phototactic response away from light, preferring the dark chamber.
To investigate the effect of humidity of woodlouse movement:
Place damp filter paper in one side of the base and a drying agent on the other side. Do not cover the lid with paper.
Place ten woodlice on the mesh in the centre of the chamber and cover with a lid.
Wait ten minutes, then take off the lid and count the number of woodlice on each side of the chamber.
Carefully move the woodlice back to the centre and repeat the experiment.
The woodlice should display a kinetic response, preferring the damp chamber.
Receptors
Our nervous system uses receptors to detect stimuli (changes in the environment) and pass on this information to the CNS. Receptors can either be whole cells (e.g. photoreceptors are cells which are sensitive to light) or proteins molecules which are found on the cell surface membrane. Each receptor is specific to a single type of stimulus, such as light, temperature or glucose concentration. When a receptor is not stimulated, there is a charge difference between the inside and outside of the membrane and it is said to be polarised. When the receptor detects a stimulus, the permeability of its cell membrane changes which changes the charge difference (potential difference) across the membrane. If the change in potential difference is large enough (i.e. it exceeds the threshold level), it will trigger an action potential (an electrical impulse) in a sensory neuron. If the stimulus is too weak, the membrane potential won’t reach the threshold, and no action potential will occur.
We contain the following receptors in our sense organs:
Chemoreceptors - receptors which detect chemicals
Thermoreceptors - receptors which detect heat
Mechanoreceptors - receptors which detect pressure (see the Pacinian corpuscle below)
Photoreceptors - receptors which detect light (e.g. rods and cones)
Pacinian corpuscles
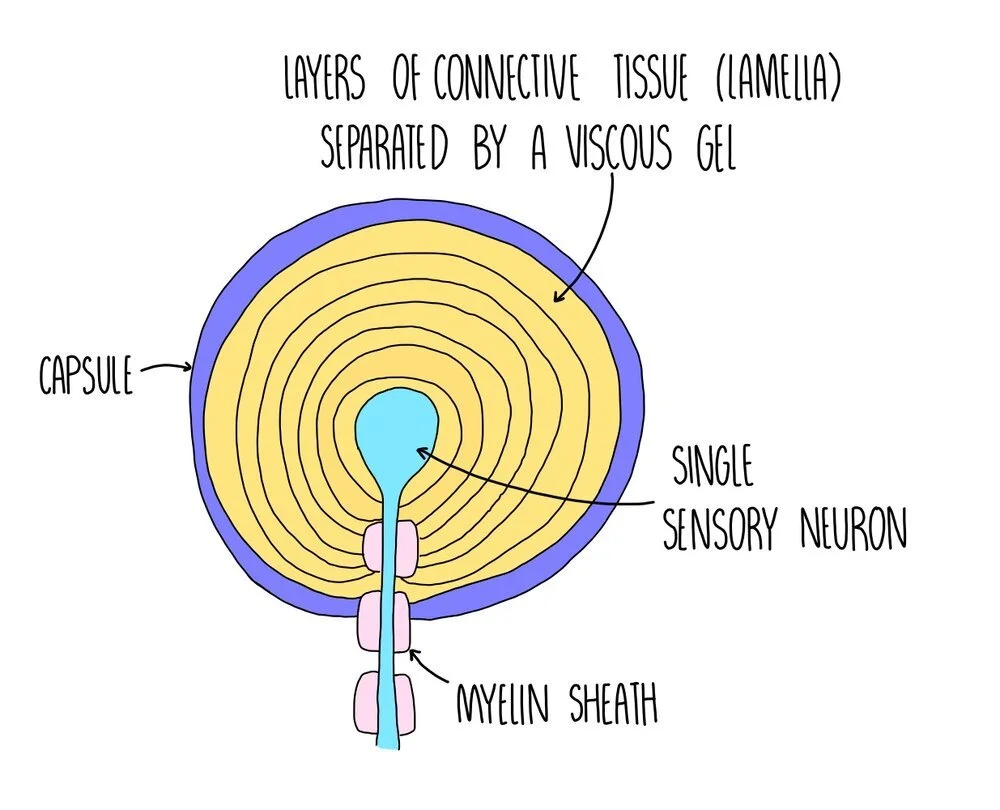
Pacinian corpuscles are receptors which respond to changes in pressure - they are a type of mechanoreceptor. They are found deep in the skin and are abundant in the feet, fingers, external genitalia and in our joints. The Pacinian corpuscle consists of a single sensory neurone, surrounded by layers of tissue which are each separated by a gel, forming an onion-like structure.
The Pacinian corpuscle contains stretch-mediated sodium ion channels in the cell surface membrane. Under normal conditions these channels are closed but when pressure is applied these channels become deformed and open, allowing a rapid influx of sodium ions. This makes the membrane potential in the neurone less negative (depolarisation), producing a generator potential which can then produce an action potential.
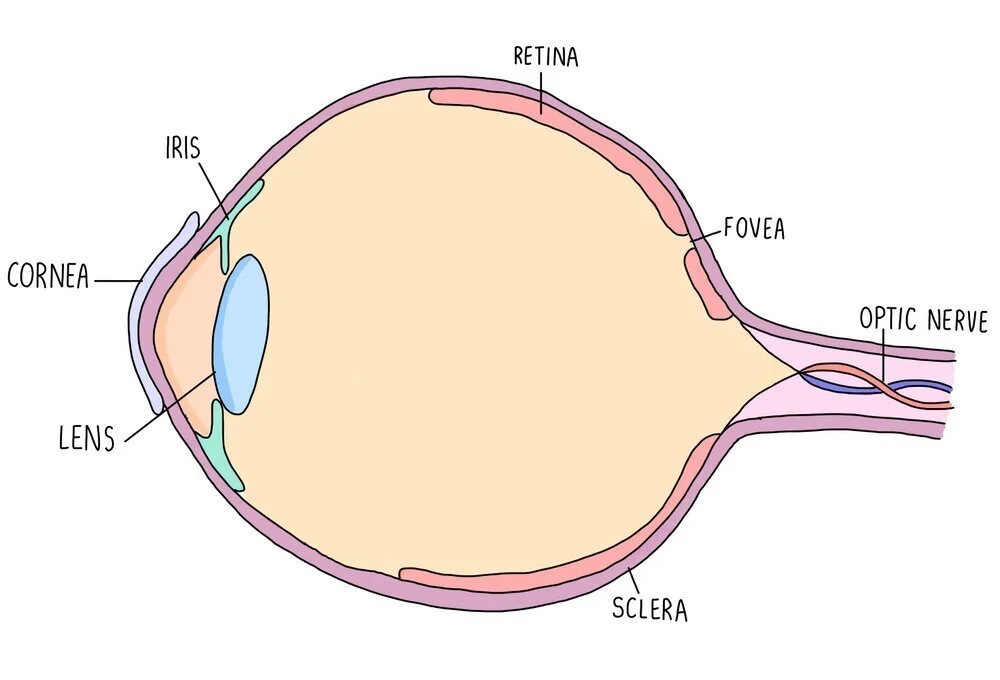
Photoreceptors
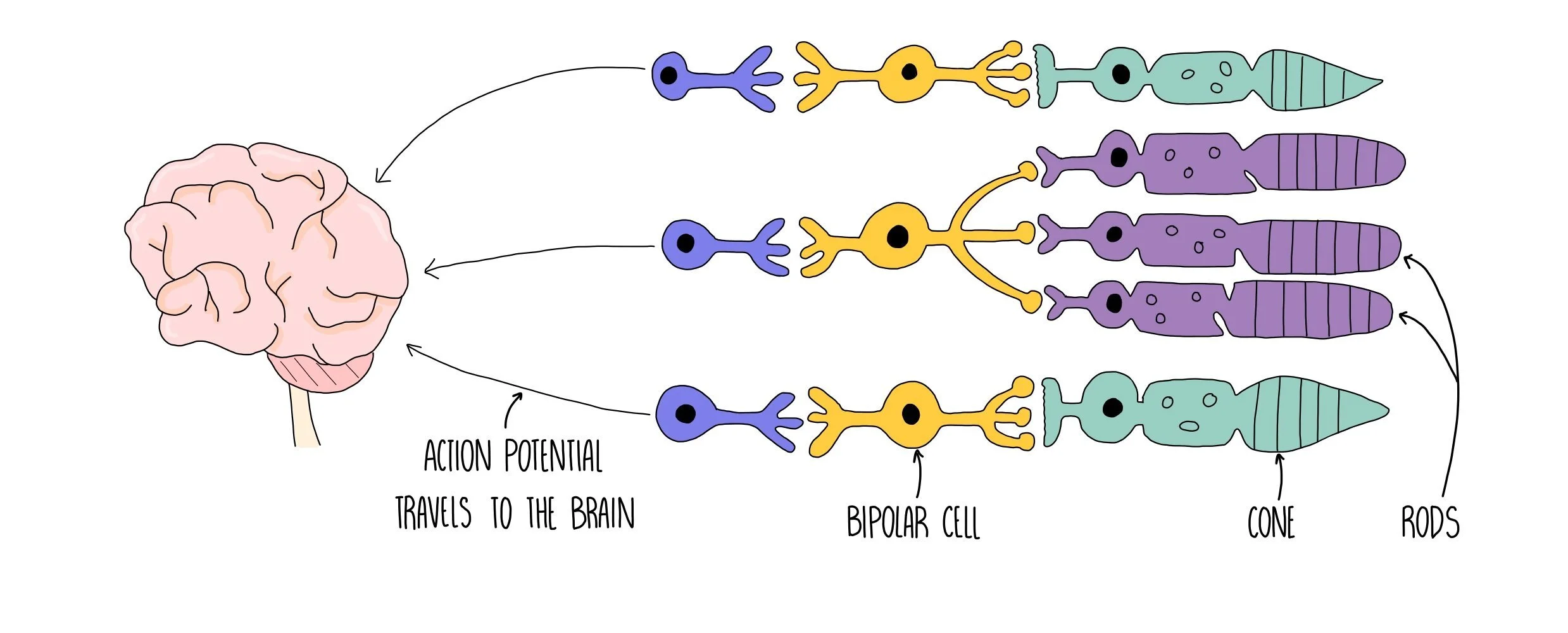
Photoreceptors are receptors which detect light and are found in the retina of the eye. There is an area of the retina called the fovea which contains a cluster of photoreceptor cells. Photoreceptors detect light as it hits the retina and send nerve impulses to the brain along the optic nerve. The region of the eye containing the optic nerve is called the blind spot since there are no photoreceptors in this region so light can’t be detected. Photoreceptors are connected to the optic nerve through a bipolar neurone.
Photoreceptors convert light into an electrical impulse:
Light hits photoreceptors and is absorbed by light-sensitive optical pigments (e.g. rhodopsin).
Light bleaches the pigments, causing a chemical reaction to take place.
This triggers the opening of sodium ion channels, which results in depolarisation of the receptor membrane.
If the membrane potential exceeds the threshold, an impulse is sent along a bipolar neuron.
The bipolar neuron passes the impulse to the brain via the optic nerve.
The human eye has two types of photoreceptors - rods and cones. Rods are mostly located along the outside of the retina while cones are clustered together in the fovea. Rods are responsible for black-and-white vision and can function in lower light levels than cones. They are much more sensitive than cones, so are the type of photoreceptor used for visualising objects in the dark. This is because many rods join one neuron, so many weak depolarisations combine to reach the threshold potential and trigger an action potential.
Cones are responsible for colour vision and are sensitive to blue, green or red light. Different cones are stimulated in different proportions, so that we see different colours. Cone cells provide good visual acuity (the ability to distinguish between two points which are close together) because each cone cell has its own synapse via a bipolar neurone which connects to the optic nerve.
Myogenic contraction
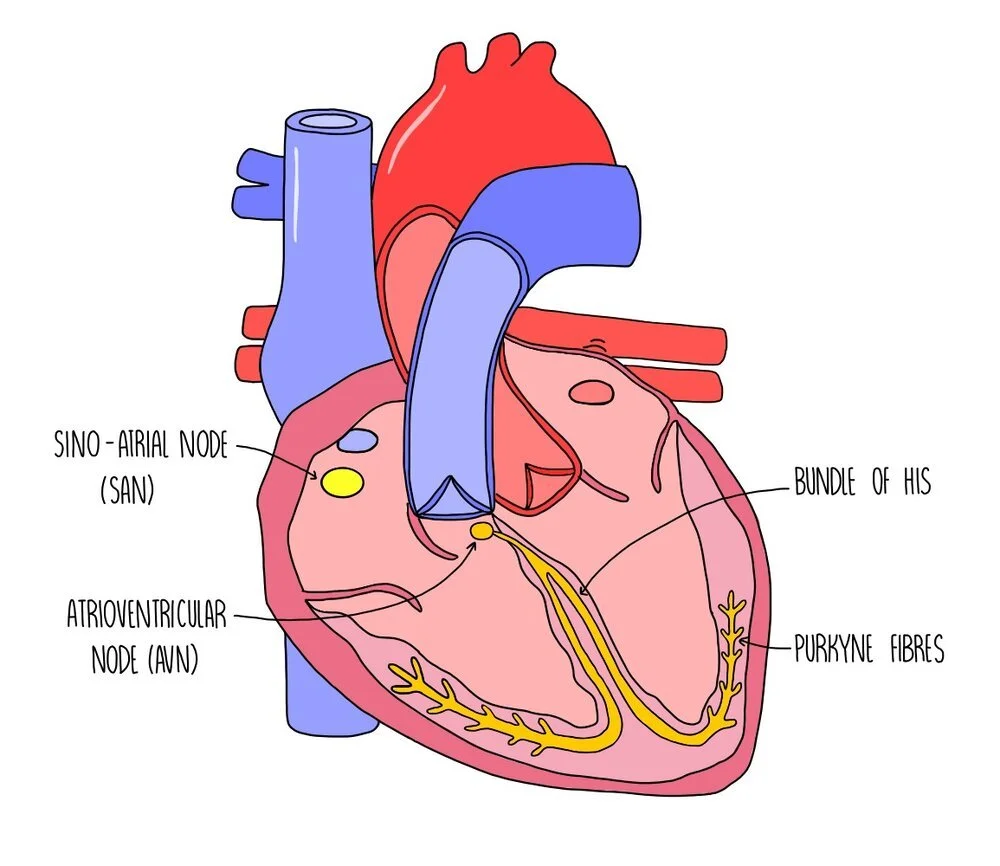
Cardiac muscle is myogenic, which means that it can contract on its own accord without being stimulated by nerve impulses. It is coordinated by a nodule in the wall of the right atrium called the sino-atrial node (SAN). The SAN is referred to as the ‘pacemaker’ of the heart because it sets the rhythm of heart contractions by creating regular waves of electrical activity which spread out to the atrial walls. This wave of electrical activity causes the right and left atria to contract at the same time. There is a band of non-conducting tissue between the atria and the ventricles which prevents the wave of electricity from reaching the ventricles. The electrical wave is instead directed towards the atrioventricular node (AVN) which is located between the atria and the ventricles. The AVN passes the wave of electrical activity onto a group of muscle fibres located in the centre of the heart, called the bundle of His. There is a delay before the wave of excitation passes onto the bundle of His to ensure that the ventricles only contract after the atria have finished emptying with blood. The bundle of His then passes on the wave of electrical activity to the Purkyne fibres, found along the bottom of the heart around the ventricular walls. This causes the left and right ventricles to contract simultaneously, from the bottom up.
Control of heart rate
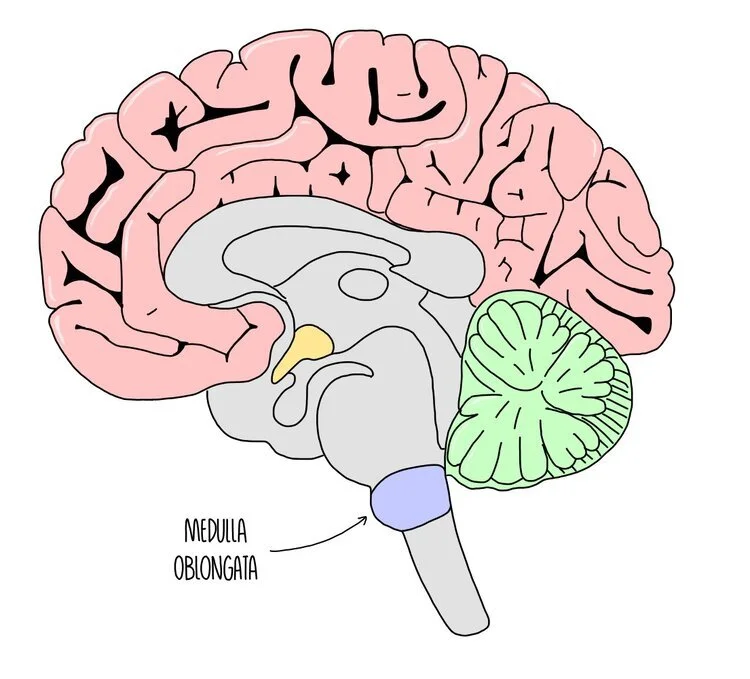
The medulla oblongata is a brain region found at the bottom of the brain, in the brain stem. It is involved in unconscious processes, such as controlling heart rate and breathing rate. A part of the medulla oblongata called the cardiovascular control centre is responsible for changing heart rate according to our body’s needs. It works by sending impulses along sympathetic or parasympathetic neurones which release different neurotransmitters onto the SAN - the SAN then modifies its rate of firing to slow down or speed up the heart rate.
Two types of receptors, baroreceptors (pressure receptors) and chemoreceptors (chemical receptors) are responsible for detecting stimuli in the blood and signalling to the medulla oblongata to modify our heart rate. Baroreceptors detect changes in blood pressure and are found in the aortic and carotid bodies. Chemoreceptors detect the concentration of oxygen in the blood. They are also sensitive to changes in pH resulting from the carbon dioxide dissolved in the blood (its reacts with the water to form carbonic acid), which is an indication of oxygen availability. Chemoreceptors are also located in the aortic and carotid bodies.
When the medulla oblongata receives signals from baroreceptors and chemoreceptors, it changes the rate at which the sino-atrial node (SAN) fires. If blood pressure or oxygen concentration is low, the cardiovascular control centre increases the rate of SAN firing through activation of the sympathetic nervous system. The sympathetic nervous system is involved in the ‘fight and flight’ response and increases heart rate through the release of a neurotransmitter called noradrenaline, which binds to receptors on the SAN. On the other hand, high blood pressure and high oxygen concentration causes the cardiovascular control centre to reduce the rate of SAN firing by activating the parasympathetic nervous system. This is involved in the ‘rest and digest’ response and decreases heart rate through the release of another neurotransmitter called acetylcholine. Acetylcholine binds to receptors on the SAN to slow down its rate of firing.
