Transport in Plants
The need for transport systems
Plants need water for photosynthesis, chemical reactions and to keep cells turgid. They also need sugars for respiration and to build organic molecules. Just like animals, they need to get rid of waste substances.
Plants can’t obtain these things by simple diffusion with their environment because they are multicellular and have a large surface area: volume ratio. Diffusion would take place too slowly so plants have mass transport systems.
Vascular bundles
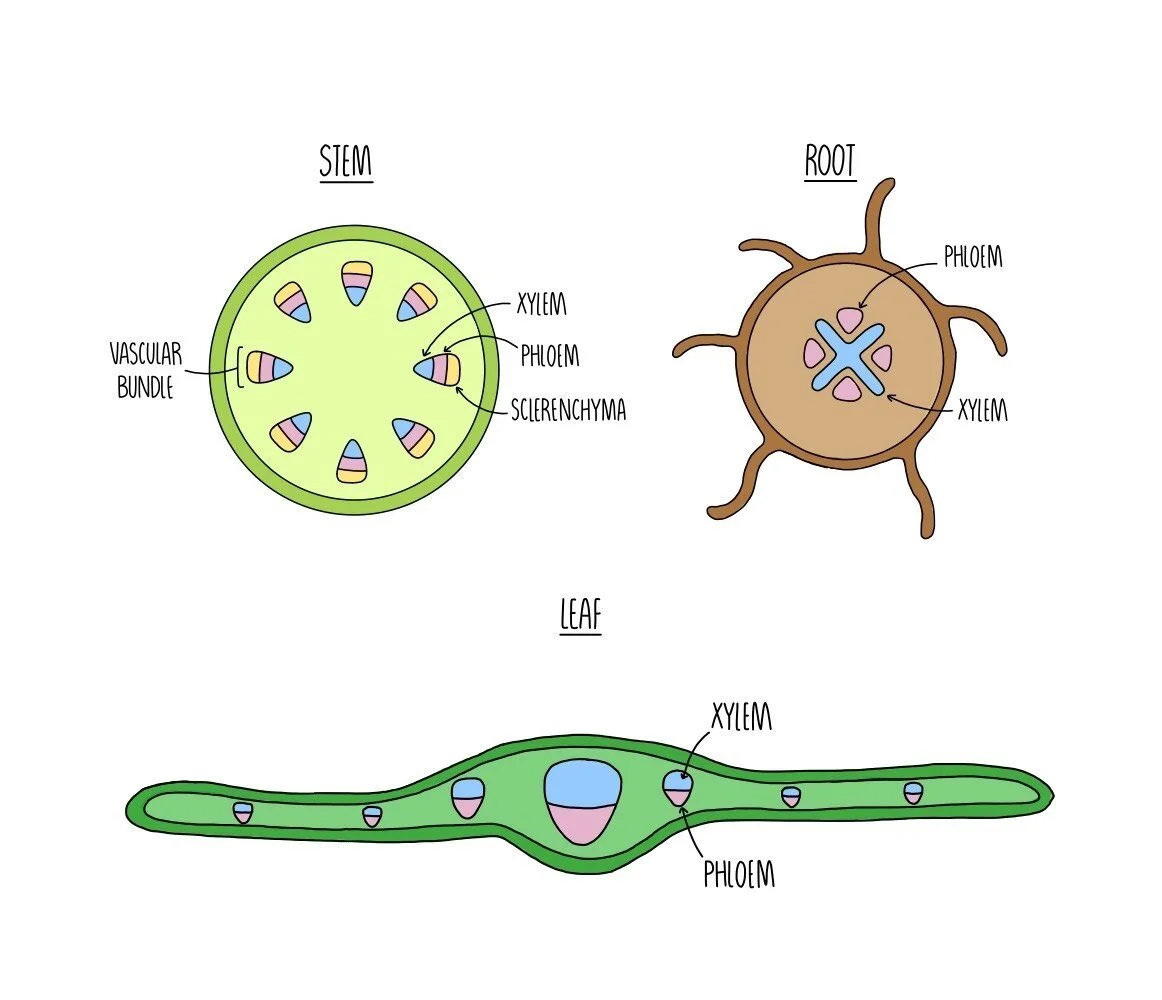
Plants contain vessels which function to transport water and sugars from one part of the plant to another. Xylem vessels transport water and dissolved mineral ions from the roots to the rest of the plant and also provide structural support. Phloem vessels transport dissolved substances, such as sucrose and amino acids, from the leaves to the rest of the plant. Xylem and phloem vessels are grouped together within the plant stem and form vascular bundles. Sclerenchyma fibres are also found within vascular bundles and provide support to the stem.
Within the plant stem, xylem vessels are found right on the inside. Phloem tissue is located in the middle of the vascular bundle and sclerenchyma fibres are found on the outside. Having the stronger xylem vessels in the centre provides strength to the stem and acts like an internal ‘scaffolding’ to support the stem and prevent it from bending in the wind.
In the root, the xylem forms a cross-like structure in the centre which is surrounded by phloem vessels. This arrangement adds strength to the root as it pushed through the soil.
Within the leaf, the xylem vessels are found towards the top of the vascular bundle with the phloem vessels found underneath.

Xylem
Xylem vessels transport water and mineral ions from the roots to the rest of the plant. They are made up of dead, hollow cells with no end cell walls. This forms one continuous tube when the xylem cells are stacked on top of each other.
The cells have no organelles or cytoplasm, which creates more space inside the vessel for transporting water. The cell walls contain pits which allow water and mineral ions to move into and out of the vessel.
The cell wall also contains a tough, woody substance called lignin, which strengthens the xylem vessel and provides structure and support to the plant.
Phloem
Phloem vessels transport dissolved substances, such as sucrose and amino acids from parts of the plant where they are made (sources) to the parts of the plant where they are used (sinks). Leaves are sources because they produce glucose from photosynthesis and parts of the plant where sugar is stored, such as roots and bulbs, act as sinks.
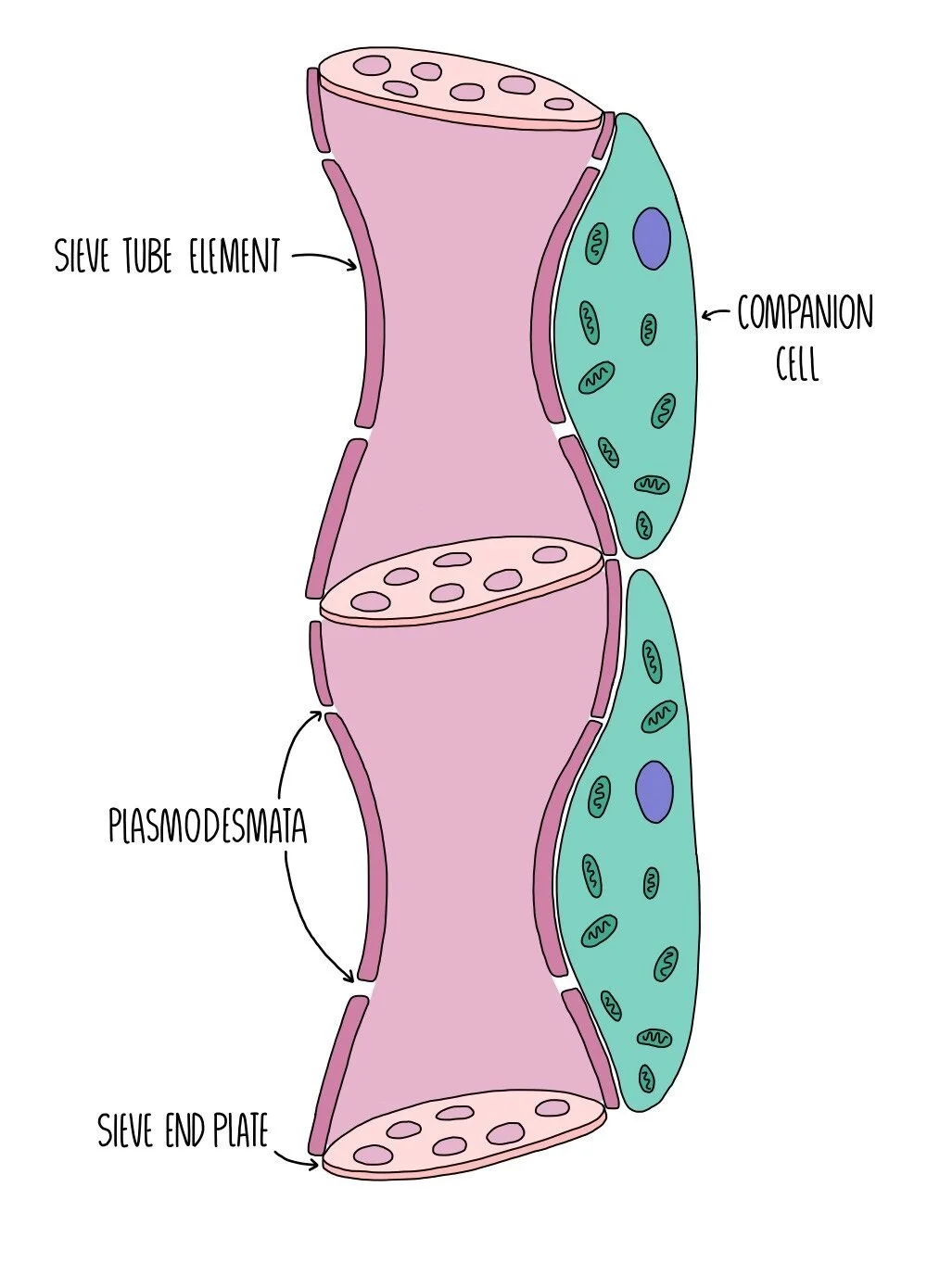
Phloem vessels are made up of two types of cell - sieve tube elements and companion cells. The sieve tube elements are living cells and are joined end-to-end to form sieve tubes. The ends of each cell consist of a ‘sieve plate’ which contains lots of holes to allow solutes to move from one cell to the next. The sieve tube cells contain no organelles and very little cytoplasm to create more space for solutes to be transported. The absence of a nucleus and other organelles means that these cells cannot survive on their own, so each sieve tube element is associated with a companion cell, which contains a nucleus and is packed full of mitochondria. The mitochondria provide lots of energy for the active loading of sucrose into the sieve tube element. The sieve tube element and the companion cell are connected through plasmodesmata (channels in the cell wall) which allow the two cells to communicate.
Dissecting plant stems
To view vascular bundles under the microscope, you first need to dissect the plant stem and prepare a tissue sample. You would do this by following the method below:

Vascular bundle in a clover leaf viewed under the microscope. Image credit: Berkshire Community College Bioscience Image Library.
Cut a thin section of the plant stem using a scalpel. Take care when using the sample and remember to cut away from you.
Place the tissue sample into water to prevent it from drying out.
Place the tissue sample into a small dish containing the stain. A common stain that is used to view vascular bundles is toluidine blue O (TBO) which stains lignin blue/green which will enable you to visualise the xylem and sclerenchyma fibres. The phloem cells and remaining tissue will appear a pink/purple colour.
Rinse the tissue samples in water and place each one onto a microscope slide.
Movement of water into roots
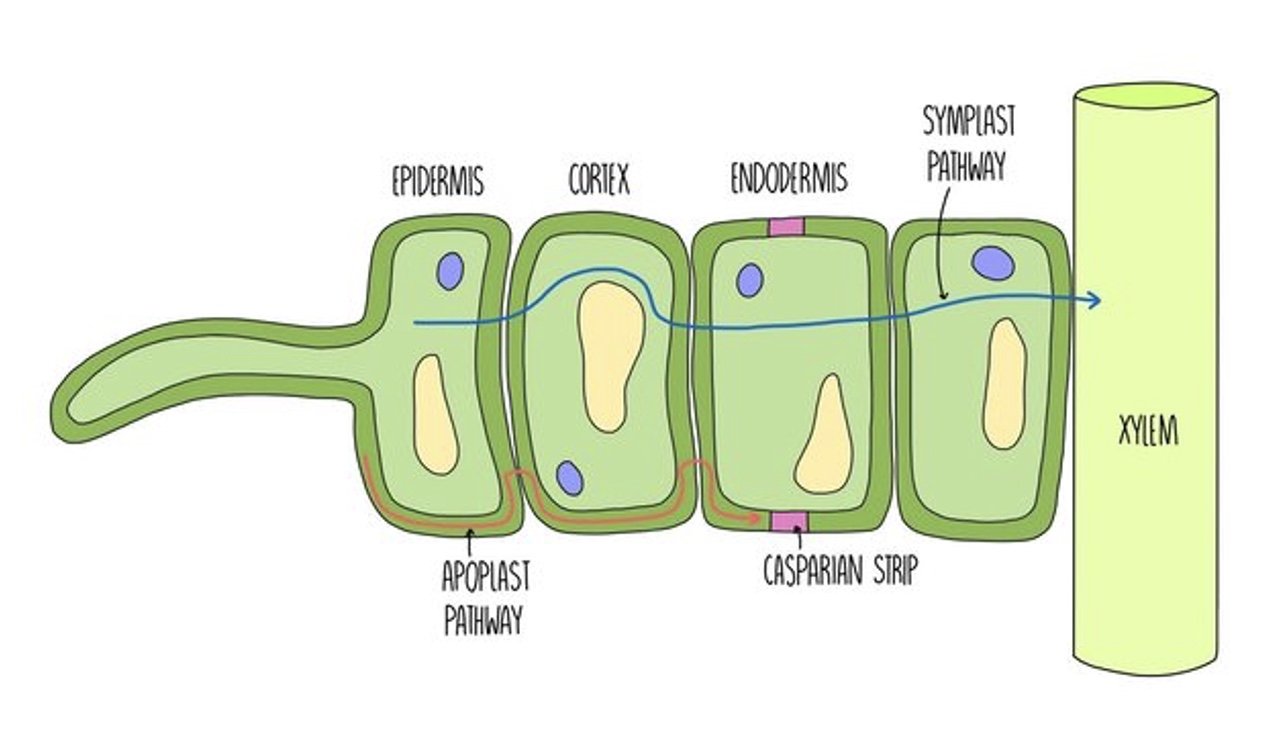
Water moves from the soil into root hair cells by osmosis. There will always be a higher concentration (or water potential) in the soil compared to the plant because water is constantly being lost through the leaves in transpiration. Water moves down its concentration gradient into root hair cells then travels through the root cortex and the endodermis before reaching the xylem.
Water can get into xylem vessels by two routes - the symplast pathway and the apoplast pathway. If water travels via the symplast pathway then it travels from cell to cell through the cytoplasm. Neighbouring cells are connected by small channels in the cell walls called plasmodesmata. However, if water is travelling via the apoplast pathway then it passes from cell to cell through the cell walls. Plant cell walls are very absorbent and water can travel through them easily, making this the main pathway that water uses to move from the roots to the xylem.
The problem with the apoplast pathway, however, is that the water (and the substances dissolved in it) bypasses the cell membrane, which controls what is going into the cell and prevents entry of toxic substances. The root, therefore, has something called the Casparian strip which is a waxy strip within the cell walls which is impermeable to water. This forces the water to go through a cell membrane which can then control which substances are allowed to enter the plant. Once water has passed through the Casparian strip, it can then reach the xylem.
Movement through xylem
Once water is in the xylem, it travels upwards - against the force of gravity - towards the rest of the plant. Water is able to move against gravity due to two forces: tension and cohesion. Tension is a ‘sucking force’ which is created when water evaporates from leaves (transpiration), pulling more water into the leaf. You can think of this in the same way as drinking water through the straw - any water which is lost from the top of the straw (as you drink) is immediately replaced with water molecules below it. Cohesion describes how water molecules are attracted towards each other. The strong hydrogen bonds between water molecules cause them to ‘stick’ together, creating a column of water. This means that when tension pulls water up the xylem, the whole column of water moves upwards.
Another force which facilitates the movement of water up the xylem is adhesion. Adhesion describes the attraction of water to non-water molecules (such as the molecules which make up the xylem walls). The attraction of water to the walls of the xylem helps water to rise up through the vessel.
Transpiration
Transpiration is the loss of water vapour through evaporation from a plant’s surface. It mainly happens through gaps in the leaf called the stomata, which need to open during the daytime to allow gas exchange. Plants need to take in carbon dioxide for photosynthesis and get rid of oxygen, which happens through the stomata. A side-effect of this is that water vapour can also diffuse out of the leaf through the stomata - this is known as transpiration. Plants will close their stomata at night (because they are not photosynthesising so gas exchange does not need to take place) which minimises transpiration.
The following factors affect the rate of transpiration:
Light intensity - increasing light intensity increases the rate of transpiration. This is because the plants are photosynthesising more and so more gas exchange needs to take place. Plants will have their stomata open for a much longer period on summer days with lots of sunlight, compared to the short winter days.
Temperature - increasing temperature increases the rate of transpiration because more heat energy means that the water molecules have more kinetic energy and will diffuse faster out of the stomata. This increases the water potential gradient between the inside and outside of the leaf.
Wind - windier conditions increases the rate of transpiration. This is because the wind will immediately blow away any water molecules that have just diffuses out of the leaf and increase the water potential gradient between the inside of the leaf and the outside.
Humidity - more humid conditions decreases the rate of transpiration. Humidity is a measure of the level of moisture in the air. The more humid the air surrounding a leaf, the lower the water potential gradient between the inside of the leaf and the outside.
Measuring transpiration rate using a potometer
You can investigate the rate of transpiration using a piece of apparatus called a potometer. A potometer actually measures the amount of water uptake by a plant. The assumption is that the amount of water uptake by a plant is equivalent to the amount of water lost through transpiration - this is not completely true since some water will be used to keep cells turgid, for photosynthesis and other chemical reactions and some water will be created from aerobic respiration. However, it is a good way of estimating the amount of transpiration taking place. You can carry out the experiment using the following method:
Take a plant and cut off a shoot. You need to do this underwater to prevent air from entering the xylem. You should also cut the shoot at a slant to increase the amount of surface area for water uptake.
Keeping the shoot underwater, insert it into the potometer.
Take the potometer out of the water, but ensure that the capillary tube is submerged in a beaker of water.
Dry the leaves and leave the apparatus for 30 mins to enable the shoot to acclimatise.
Ensure that all other variables are controlled - e.g. light intensity and humidity.
Shut the tap so that the capillary tube system is closed them remove the end of the capillary tube from the beaker of water until one air bubble has formed. Place the capillary tube back into the beaker of water.
Record the starting position of the air bubble then time how long it takes for the air bubble to move along the capillary tube. Record the distance moved by the bubble and calculate the speed (distance moved per unit time).
The speed of the bubble can be used to estimate the rate of transpiration of the plant shoot.
Xerophytes
Xerophytes are plants which are adapted to living in regions where water is scarce. Examples include cacti, pineapple and marram grass. They have a number of adaptations which enables them to survive in such harsh conditions:

Waxy layer on the epidermis - this waterproof outer layer reduces evaporation from the surface because water cannot easily pass through (it is impermeable).
Sunken stomata - xerophytes have stomata which are sunken in pits. The pits shelter the stomata from the wind, reducing the water potential gradient between the inside of the leaf and the outside.
Hairs on epidermis - hairs on the epidermis trap water vapour, reducing the water potential gradient between the inside and outside of the leaf.
Spines - many xerophytes have spines instead of leaves which reduces the surface area for water loss.
Rolled leaves - curled leaves traps water vapour, reducing the water potential gradient for transpiration. It also reduces the surface area of the leaf for water loss.
Closure of stomata - xerophytes can close their stomata during conditions of particularly high temperature or light intensity. This reduces transpiration at times when the rate of transpiration would be very high.
Hydrophytes
Hydrophytes are plants which live on water, such as water lilies. Because oxygen does not dissolve well in water, hydrophytes need adaptation to enable them to cope with low oxygen levels. They are adapted in the following ways:

Stomata on the upper surface - usually stomata are found on the underside of plant leaves but for hydrophytes, this side will be submerged in water. Instead, hydrophytes have their stomata on the upper surface of their floating leaves to maximise gas exchange.
Air spaces - pockets of air in the plant tissue help the plant to float and can be used to store oxygen for aerobic respiration.
Flexible leaves and stems - the flexibility of the leaves and stems helps to prevent damage from water currents. Unlike land plants, which need a sturdy stem to keep upright, hydrophytes are supported by the water around them.
Translocation
Translocation is the movement of dissolved substances, such as sucrose and amino acids, from parts of the plant where the substances are made to other parts of the plant where they’re needed. Translocation takes place in the phloem - transport vessels made up of two types of cell, sieve tube elements and companion cells. The parts of the plant which make these substances are referred to as sources (e.g. the leaves) and the parts of the plant which store or use the substances are called sinks (examples include bulbs and roots). When sucrose reaches a sink, it is converted into starch for carbohydrate storage. This maintains a concentration gradient between the source and the sink, so that more sucrose moves into the source. Translocation is an active process, so if respiration is reduced or inhibited (e.g. using a respiratory toxin), translocation will be impaired.
The Mass Flow Hypothesis
The mass flow hypothesis is a theory which attempts to explain how solutes are transported from source cells into sinks through the phloem. It isn’t concrete, but it is the best-accepted theory we currently have based on the available evidence. It states that mass flow of solutes takes place in the phloem in the following stages:
Sucrose moves from companion cells into sieve tube elements by active transport.
This reduces the water potential of the sieve tube element.
Water moves into the phloem by osmosis, which increases the hydrostatic pressure.
There is a pressure gradient with high hydrostatic pressure near the source cell and lower hydrostatic pressure near the sink cells.
Solutes move down the pressure gradient towards the sink end of the phloem.
Solutes move into sink cells and are converted into other molecules (e.g. starch).
The removal of solutes increases the water potential at the sink end, causing water to move out of the phloem by osmosis. This maintains the hydrostatic pressure gradient between the source and the sink.
Active Loading
You also need to know how sucrose is actively transported into the sieve tube elements in the first place. This process involves the companion cells which are associated with each sieve tube element:
The companion cell actively transports hydrogen ions into the surrounding cells.
This creates a hydrogen ion gradient between the surrounding cells and the companion cell.
Hydrogen ions move back into the companion cell down their concentration gradient through a co-transporter protein.
Whenever a hydrogen ion moves through the co-transporter, a sucrose molecule is also transported into the companion cell, against its concentration gradient.
The same process occurs to transport sucrose from the companion cell into the sieve tube element.